В статье отмечено, что беспозвоночные животные являются важной и малоизученной составляющей полупустынных экосистем Казахстана. Данные по энтомофауне, а также другим группам традиционно используют для биоиндикации антропогенного и техногенного загрязнения. Фауна беспозвоночных района Северного Прибалхашья изучалась в связи с отчуждением земель для восстановления деятельности промышленного комплекса. Авторы определяют, что в качестве неизмененного природного ландшафта была выделена буферная зона. Исследование герпетобионтов на уровне крупных таксонов показало зависимость распределения беспозвоночных от экологического состояния биоценозов. Выполнен анализ динамической плотности и трофической структуры беспозвоночных.
В настоящее время к герпетобионтам относят напочвенных (подстилочных) беспозвоночных животных из надкласса многоножки, классов ракообразные, паукообразные и открыточелюстные насекомые, населяющих поверхность почвы. Ведущую роль в герпетобии играют открыточелюстные насекомые, среди которых преобладают представители отрядов жесткокрылые (Coleoptera), перепончатокрылые (Hymenoptera) и клопы (Hemiptera).
Известно все более широкое использование почвенных жесткокрылых, в частности жужелиц (Co^ptera, Carabidae), как количественных и качественных индикаторов биоценозов. Они могут служить точными, надежными и удобными биологическими маркерами механического состава почв, солевого и гидротермического режимов, типа почв, их рельефа, растительности. Важна роль жесткокрылых как показателей изменения биоценозов при антропогенных воздействиях: распашке, выпасах, полезащитном лесоразведении, мелиоративных мероприятиях [1, 2].
Исследование антропогенной трансформации экосистем, выявление видов-индикаторов этого процесса весьма актуальны в экосистемах сухих степей, полупустынь и пустынь.
Значительные территории Республики Казахстан все интенсивнее вовлекаются в сферу антропогенного освоения, неизбежным следствием которого является ухудшение экологической обстановки региона. На сегодняшний день одной из актуальных экологических проблем является загрязнение окружающей среды в районах действия золотодобывающих и других горно-обогатительных заводов [3]. Рост антропогенной нагрузки при освоении месторождений золота приводит к изменению химических и физико-химических свойств почвы, нарушению гидрологического режима территорий, потере биологического разнообразия. Особенно сильно воздействуют стрессовые техногенные факторы на неустойчивые и хрупкие пустынные экосистемы, среди которых животные и растительные сообщества Северного Прибалхашья. Однако трансформация флоры и фауны региона изучена мало.
Северное Прибалхашье отличается большой засушливостью и континентальностью, в связи с чем воздействие стрессовых антропогенных факторов проявляется здесь с большей силой [3]. Формирование уникального фаунистического комплекса беспозвоночных региона исторически обусловлено взаимодействием основных факторов: глубоким внутриконтинентальным положением региона, режимом влагообеспеченности за счет близости озера Балхаш и связанных с ним водоемов, крайне неравномерным распределением атмосферных осадков, высокодифференцированным горным рельефом, создающим богатый спектр локальных экологических условий.
В районе Северного Прибалхашья расположены многочисленные предприятия горнодобывающей промышленности. Для них характерно интенсивное воздействие на окружающую природную среду, неизбежно вызывающее ее изменение. В процессе производства нарушается полностью или частично сложившееся экологическое равновесие в зонах размещения промышленных объектов (шахт, рудников, обогатительных фабрик). Эти изменения проявляются в отчуждении территорий для производства горных работ, истощении и загрязнении подземных и поверхностных вод, затоплении и заболачивании подработанных территорий, обезвоживании и засолении почв, загрязнении вредными веществами и химическими элементами атмосферного воздуха, изменении микроклимата. В связи с этим все большее внимание уделяется вопросам создания экономически обоснованных и экологически безопасных горнодобывающих предприятий. Особую актуальность приобретают вопросы практического изучения фауны «культурного ландшафта» и исследования перестройки животного населения, которые происходят при широком освоении новых земель и их природных ресурсов [4].
При оценке качества природной среды и ее трансформации в результате техногенного воздействия рассматривается такие важные характеристики, как состояние фауны почв, структура комплекса герпетобионтных беспозвоночных и устойчивость его к техногенному воздействию, возможный уровень стабилизации процессов деградации фауны.
Конечная цель биоиндикационных работ — создание оптимального ландшафта, под которым понимается такая форма организации территории, которая позволяет не только обеспечить максимальную продуктивность земельных угодий, но и удовлетворить рекреационные, эстетические, санитарно-гигиенические и другие требования к природной среде.
Еще в 1960 г. началась систематическая разведка группы месторождений «Пустынное» в Акто-гайском районе, в 120 километрах от города Балхаша. В 1998 г. выполнялась разработка оксидных руд месторождения для извлечения золота. В силу выбранной технологии потенциал месторождения не был реализован полностью, и после длительного перерыва к реконструкции горно-металлургического предприятия «Пустынное» приступили в 2012 г. Для оценки состояния природных комплексов были начаты флористические и фаунистические исследования. В качестве удобных объектов проведения биомониторинга и биоиндикационных работ были выбраны герпетобионты. Целью нашей работы было изучение фауны наземных беспозвоночных на участках различной степени техногенного воздействия и выявление возможных индикаторных видов. Актуальность изучения этих животных диктуется их обилием и важной ролью в биоценозах, чуткостью к изменениям природных режимов.
Материалы и методы исследования
Полевые работы проводились в период осени и весны 2013 г. на территории золотодобывающего месторождения «Пустынное» в зоне боялычевых пустынь. На исследуемой территории обычны бело-земельно-полынно-боялычевые, тырсиково-белоземельно-полынные сообщества. Были выделены экспериментальные участки в трех зонах, отличных по степени воздействия: буферная — 3 км от техногенного источника, санитарная — 1 км от техногенного источника и собственно промышленная, расположенная по розе ветров.
Буферная зона представляет собой холмистую территорию. Растительность злаково-кустарнико-вая, проективное покрытие 80-90 %. Встречаются боялыч, лебеда седая, тасбиюргун, курчавка, ковыль. Обилен ревень татарский.
Санитарная зона имеет растительный покров мезоксерофильного типа, в понижениях растет больше злаков. Встречаются разнообразные луковые и лилейные, ложноочитки. Для этой зоны характерно обилие лебеды седой и ферулы татарской. Проективное покрытие 70-80 %.
Для промышленной зоны характерна значительно большая изреженность растительности. Проективное покрытие 55-65 %. Почва сильно уплотнена. Многие растения здесь представлены более низкорослыми формами (ревень татарский и ферула татарская). Почти отсутствуют тюльпаны. На солонцеватых щебнистых почвах растет гультемия персидская, местами образующая заросли.
Герпетобионтные беспозвоночные, активно передвигающиеся по поверхности почвы, регистрировались в течение суток при помощи ловушек и визуального учета. Также учитывались обитатели укрытий, щелей и трещин верхнего слоя почвы. Всего за время проведения исследований было поставлено 202 ловушки. Обработано и определено 547 экземпляров беспозвоночных.
Для сбора беспозвоночных применялись традиционные энтомологические методики: лов в почвенные ловушки Барбера, сбор на свет и сбор одиночных насекомых с земли и лов на лету с помощью стандартного энтомологического сачка.
Ловушки Барбера использовались для ловли ползающих по поверхности почвы насекомых. Метод ловушек позволяет учитывать динамическую плотность, т.е. число особей, пересекающих в единицу времени линию определенной длины [5, 6]. Определялась динамическая плотность (улови-стость) беспозвоночных за время, на которое устанавливались ловушки.
Светоловушки были эффективны при сборе ночных насекомых. Маршрутный учет использовался при поиске крупных беспозвоночных, выходящих на дневную поверхность в определенные часы суток в местах с разреженной растительностью, для изучения суточной и сезонной активности животных в биотопе. Учитывались животные, встреченные на маршруте 1,5 км. Ширина учетной полосы выбиралась в зависимости от того, насколько объект учета заметен. На открытых участках применялся учет беспозвоночных под камнями (и другими укрытиями). Собранные беспозвоночные учитывались в общем списке видов. Идентификация групп проводилась по соответствующим определителям фауны Казахстана и сопредельных стран [7, 8].
Результаты и их обсуждение
Герпетобионтная фауна беспозвоночных боялычево-полынных биоценозов Северного Прибалхашья достаточно разнообразна. В первой половине сентября, согласно нашим исследованиям, основное ядро составляют представители отряда двукрылых (семейство Myscidae — настоящие мухи), жесткокрылых (главным образом семейства Tenebrionidae — чернотелки и Carabidae — жужелицы), чешуекрылых (семейства Pterophoridae — пальцекрылки, Puralididae — огневки, Noctuidae — совки), прямокрылых (семейства Acrididae — саранчовые настоящие и Gryllidae — сверчки настоящие)(рис. 1).
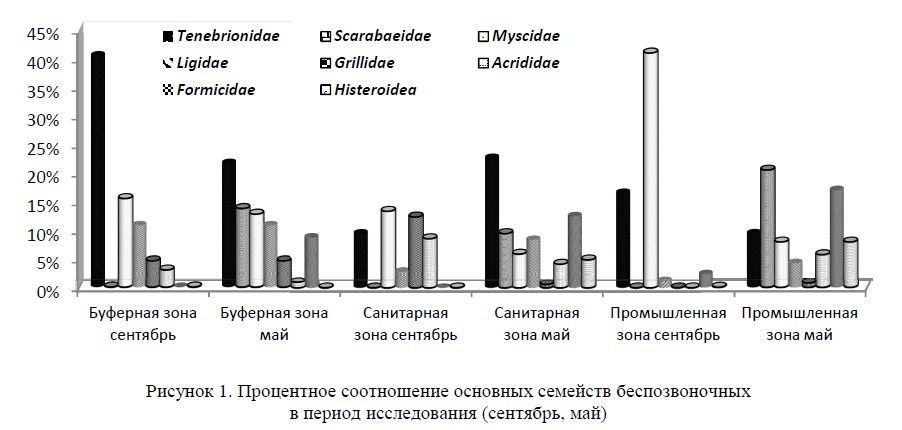
В первой половине мая основное ядро составляют представители отряда жесткокрылых (главным образом семейства Tenebrionidae — чернотелки и Scarabeidae — пластинчатоусые), перепончатокрылых (семейство Formicidae — муравьи), двукрылых (семейство Myscidae — настоящие мухи).
Обитающие здесь фитофилы в своем развитии и питании связаны с растениями-доминантами: боялычем, полынью, тамариксом и др. Популяции их немногочисленны, но в отдельные годы разви-тся обильно (например, боялышевая совка — Odontelia arbusculae Sukh.).
Из саранчовых характерны: Calliptamus italicus L. — итальянский прус, и личинки, и имаго его сильно вредят пастбищам и сенокосным угодьям; Dociostaurus brenicollis — крестовичка, пустынно-степной вид; Dociostaurus kraussi Ing. — атбасарская крестовичка, пустынно-степной обычный вид. Место обитания — полупустынные участки с засоленными почвами и полынной или злаково-эфеме-роидной растительностью. Предпочитает злаки.
Из других ортоптероидных насекомых на местообитаниях этого биоценоза отмечены представители рода Gryllomorpha sp. (Gryllidae).
За счет ортоптероидных насекомых живет большая ^упга хищных (скутигеры, тарантулы, пауки-скакуны, богомолы из рода Mantis, Bolivaria, Rivetina, мухи-ктыри и др.) и паразитических насекомых. Из энтомофагов часто встречались перепончатокрылые, особенно Prionyx subfuscatus Dahl. (Sphecidae), обычный на сухих лугах и в пустынной зоне. Охотится на саранчовых. В наших сборах преобладали эндопаразитические наездники подсемейств Ichneumoninae, Cryptinae, осы сем. Scoliidae (эктопаразиты личинок пластинчатоусых, долгоносиков) и мухи из сем. Bombylidae, паразитирующие в кубышках саранчовых. На теле саранчовых паразитируют клещи краснотелки (сем. Trombidiidae). Осы Bembix rostrata L. (Crabronidae) своих личинок кормят убитыми крупными мухами (Tabanidae, Syrphidae), а дорожные осы Pompilus viaticus L. (Роmpilidae) — охотники на крупных пауков. Роющие осы охотятся на мелких беспозвоночных.
Под камнями среди остатков растительности встречаются мокрицы из рода Hemilepistus, скорпионы, многоножки, паразитические клещи, пауки, тарантулы из рода Lycosa. Среди кустов боялыча и курчавки пауки сем. Araneidae (Argiope lobata Aud и др.) строят колесовидные сети и ловят крупных летающих насекомых.
Самая большая группа насекомых— жесткокрылые, составляющая, по нашим данным, 42 % фауны герпетобионтов-беспозвоночных. Наиболее широко в период исследования были представлены чернотелки и пластинчатоусые.
Жужелицы (Carabidae) также встречаются довольно часто. Хорошо летят на свет мелкие жужелицы Bembidion quadripustulatus Serv. Для середины сентября типичен Cymindis picta Pal., пустынная жужелица, не проникающая в более умеренные полосы. Норы грызунов заселяет Pseudotaphoxenus rufitarsis (Fischer) — псевдотафоксенус рыжепалый.
Одним из крупнейших семейств в данном биоценозе и вообще в пустыне является семейство чернотелок. Многие из них отрицательно воздействуют на растения-эдификаторы. Чернотелки встречаются с ранней весны и до глубокой осени, активны в сумерках и в утренние часы. Зимуют взрослые. Летят на свет.
На участках с уплотненными щебнистыми почвами отмечен хищный Tentyria nomas Pall. — чернотелка бродячая.
Любят песчаные места и яркое солнечное освещение виды Pimelia interpunctata Klug — толстяк внутриточечный и Pimelia cephalotes — толстяк головастый. Относятся к псаммофилам и отличаются хорошо развитыми на ногах щетками из длинных волосков, которые облегчают передвижение по песку и закапывание в него. Их личинки повреждают корни эфемеров, полыни, однолетних и многолетних солянок.
В ранневесенний период появляется Adesmia anomala (F.-W.) — адесмия непривычная, исключительно обитатель пустынь, в связи с чем имеет необычайно длинные ноги. Встречается до глубокой осени. Обитает во всех биоценозах, предпочитает уплотненные щебнистые почвы, но селится и на закрепленных песках. Днем жуки скрываются в своих норках или в норах песчанок. Отмечаются в основном на поверхности почвы, питаются отрастающими растениями, личинки живут в почве у корней полыни, кейреука, боялыча, осоки и поедают всходы.
Жуки подсемейства Elaterinae, род Blaps, зимуют в почве и растительной подстилке. Основными местами зимовок служат многолетние травы. Жуки активны в утренние и вечерние часы, днем прячутся в укрытия, например под растительными остатками. Питаются преимущественно увядшими растениями. Предпочитают участки с редкой растительностью, хорошо прогреваемые солнцем. Сухолюбивы, влажных почв не выносят. Личинки питаются семенами и подземными частями растений. Многочисленны также виды песчаных чернотелок Blaps pruinosa Fald. и Blaps deplanata Men.
Эвритопный ксерофил, предпочитающий сухие участки, Opatrum sabulosum L., относится к мно-гоядным вредителям. Жуки повреждают весной всходы злаков, у проросших растений объедают семядоли и молодые листочки около поверхности почвы, подгрызают и перегрызают стебельки молодых растений у основания. Личинки повреждают семена, корни, подземные части стеблей различных культур.
Многочислен в боялычевом биоценозе видовой состав чешуекрылых: семейства огневок (Pyralidae), пальцекрылок (Pterophoridae), волнянок (Limantriidae), пядениц (Geometridae). Связано с обитанием в почве сем. Noctuidae. Бабочки пустынной совки (Aleucamtis flexuosa Men.) в большом количестве летят на свет осенью.
Количество семейств, зарегистрированных весной и осенью, в буферной зоне — 7, в санитарной — 11, в промышленной зоне — 5.
Исследования фауны наземных беспозвоночных на участках различной степени антропогенного воздействия в сентябре показали, что доля жесткокрылых (14,4 %) минимальна в санитарной зоне (промежуточной по антропогенному воздействию), а в промышленной зоне численность жуков повышается до 29 %. В сентябре везде основным семейством остаются Tenebrionidae.
Исследования в мае выявили, что доля жесткокрылых меняется не столь значительно — от 38 % (в буферной зоне) до 44 % (в санитарной зоне) и 42 % (в промышленной зоне). Уловистость чернотелок меняется от 0,36 экз. на ловушко/сутки (в промышленной зоне) до 1,1 экз. на ловушко/сутки (в санитарной зоне) и 0,6 экз. на ловушко/сутки (в буферной зоне). Определено 12 видов чернотелок.
С осени до весны на буферной территории чаще, чем на других участках встречались пустынные мокрицы U = 0,2, свидетельствуя о большем разнообразии микроусловий, в том числе создаваемых растительностью.
Однако максимальная уловистость ловушек осенью и весной характерна для санитарной территории (U = 2,9 и 4,6 экз. на ловушко/сутки). Наибольший вклад здесь вносят группы Tenebrionidae (жуки-чернотелки/жуки) U = 0,3/0,4 экз. на ловушко/сутки; Diptera (двукрылые) U = 0,6 экз. на ловушко/сутки; Hymenoptera (перепончатокрылые: хищные роющие, дорожные и песочные осы) U = 0,3 экз. на ловушко/сутки; Orthoptera (прямокрылые) U = 0,6 экз. на ловушко/сутки и Lepidoptera (бабочки, в основном фитофаги-огневки) U = 0,75 экз. на ловушко/сутки (рис. 2).
Однако чернотелки здесь представлены крайне однородно, только видами р. Blaps. А среди двукрылых (7 семейств) доминируют минирующие мухи и настоящие мухи, численность которых сильно возрастает к концу лета — началу осени, что является фенологической особенностью семейства.
Жуки-чернотелки — зональная группа аридных районов Казахстана — в биоценозах играют роль, детрито-, сапро-, фитофагов. Жуки и их личинки участвуют в почвообразовательных процессах и служат почвенными индикаторами. В исследуемых ценозах в сентябре основным семейством являются Tenebrionidae. Определено 8 фоновых видов чернотелок: Adesmia anomalia dejeani Gebler, 1841; Anatolica gibbosa (Steven, 1829); Tentyria nomas (Pallas, 1781); Cyphogenia limbata Fischer von Waldheim, 1820; Pimelia cephalotes (Pallas, 1781); Pelorocnemis punctata (Gebler, 1845); Pimelia interpunctata Schuster, 1938; Blaps halophila Fischer von Waldheim, 1832.
Уловистость чернотелок максимальна для буферной зоны, где она примерно в 2 раза выше, чем для двух других зон (жуки чернотелки/жуки) U = 0,7/0,8 экз. на ловушко/сутки (рис. 2). Для этой же территории определено максимальное количество видов (7).
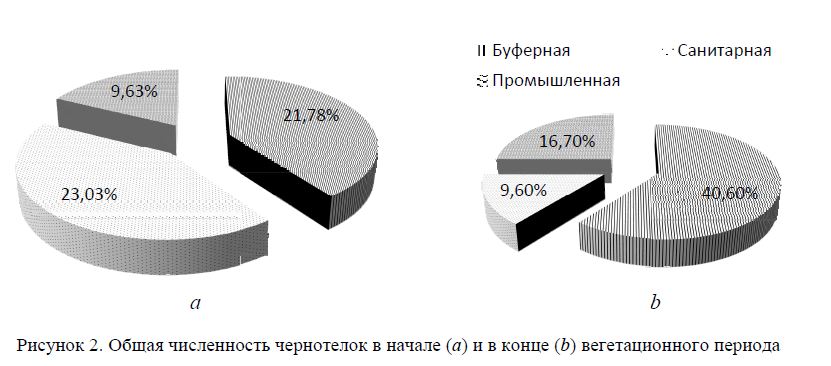
Исследования в мае выявили, что уловистость чернотелок меняется от 0,36 экз. на ловуш-ко/сутки (в промышленной зоне) до 1,1 экз. на ловушко/сутки (в санитарной зоне) и 0,6 экз. на ло-вушко/сутки (в буферной зоне).
Высокая сезонная уловистость на промышленной территории U = 2,6 и 3,8 экз. на ловушко/сутки обусловлена отличительными доминантными группами: осенью — Tenebrionidae (жуки-чернотелки) U = 0,4 и Diptera (двукрылые) U = 1,2 экз. на ловушко/сутки; весной — Scarabaeidae U = 0,78 и Hy-menoptera U = 0,67 экз. на ловушко/сутки. Однако чернотелки здесь представлены в основном видами р. Blaps.
Основная доля численности чернотелок на всех трех пробных площадках приходится на вид Anatolica gibbosa (30-45 %). Это типичный пустынный вид, явно выраженный ксерофил, избегающий мест с повышенной влажностью. Малочувствителен к внешним воздействиям.
В буферной и промышленной зонах в значительном количестве был представлен Blaps halophila (27.3 % и 23 % соответственно). Жуки активны в утренние и вечерние часы. Днем, избегая высокой температуры, прячутся в укрытиях. Медляк степной — крайне сухолюбивый вид, предпочитает места с редкой растительностью, хорошо прогреваемые солнцем, с высоким уровнем засоления. Численность чернотелок рода Blaps постоянна, однако вид B. halophila не был встречен в санитарной зоне площадке ни в сентябре, ни в мае, что мы связываем с повышенной влажностью почвы и обилием плотной злаковой растительности в местообитании. Эти чернотелки избегают участков с повышенной влажностью и рыхлостью почв.
Неоднородна на исследуемой территории численность чернотелки Tentyria nomas. Она возрастает от буферной зоны (4,55 %) к санитарной (10,53 %) и промышленной зоне (23,08 %). Особи всеядны, хищничают, отдают предпочтение некрофагии. Вид обитает на сухих, плотных почвах со скудной растительностью. Градиент численности T. nomas от буферной до промышленной зоны может свидетельствовать о постепенном изменении экологических условий, благоприятствующих обитанию вида (изрежение растительного покрова, уплотнение почвы, уменьшение межвидовой или трофической конкуренции).
Типичный обитатель пустынь региона Северного Прибалхашья — Adesmia anomalia характеризуется достаточно равномерной численностью на всех пробных участках. Численность адесмий в буферной зоне — 9,09 %, в санитарной — 5,06 %, в промышленной зоне 7,69 % (рис. 3). Ксерофильный вид. Селятся на плотных почвах и закрепленных песках.
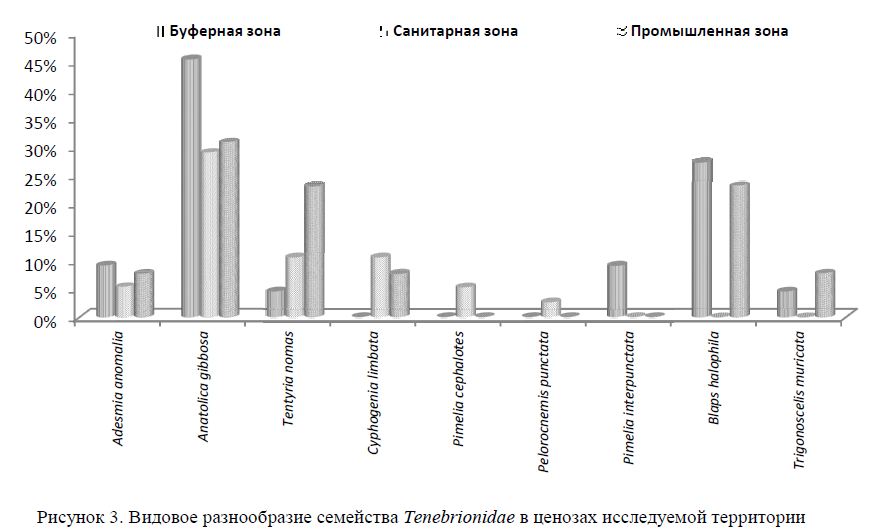
Рисунок 3. Видовое разнообразие семейства Tenebrionidae в ценозах исследуемой территории
Чернотелка Cyphogenia limbata была встречена только в санитарной и промышленной зонах (10,53 % и 7,69 %). Ксерофильный вид.
Виды Pimelia cephalotes и Pelorocnemis punctata отмечены лишь в санитарной зоне, избегали из-реженных пространств промышленной зоны. Их процент в сборах значительно ниже, чем у других видов — 5,23 % и 2,63 % соответственно. Pimelia interpunctata найден только в естественных ценозах. Эти данные согласуются с экологической характеристикой видов р. Pimelia, относящихся к мезоксе-рофилам, предпочитающих места с небольшой влажностью и рыхлой почвой, достаточным количеством растительности.
Малочислен и Trigonoscelis muricata, обнаруженный на буферной (4,55 %) и промышленной площадках (7,69 %), как и Gnaptor spinimanus.
Таким образом, мы можем заключить, что доминантной группой герпетобионтов естественных ландшафтов в период исследования является семейство Tenebrionidae (41-21 % общей численности). Фоновыми семействами являются Myscidae (16-11,6 %о) и Ligidae (11 %о). В ненарушенных ценозах наблюдается стабильность доминантных и фоновых групп по сезонам.
В условиях техногенной трансформации растительного и почвенного покрова происходит смена доминантных и фоновых семейств герпетобионтов, Доминантной группой в сентябре становятся двукрылые Muscidae (41 %); в мае — жесткокрылые Scarabeidae (20,7 %) и перепончатокрылые Formicidae (17,2 %). Сопутствующей фоновой группой становятся Tenebrionidae (16,7 % и 9,6 %). За прошедшие 50 лет со времени разведки и разработки месторождения на территории санитарной зоны сформировалось пространство экотона. Здесь выявлен краевой эффект: наблюдается максимум уловистости ловушек, встречается наибольшее количество видов, семейств и отрядов герпетобионт-ных беспозвоночных.
Кроме того, отмечено, что изменение трофической структуры комплекса герпетофауны вдоль градиента трансформации местообитаний носит характер, обратный естественному для аридной зоны: доля сапрофагов возрастает в 2,5 раза (также уменьшается доля фитофагов), доля хищников и энтомофагов увеличивается почти в 3 раза.
Поиск индикаторных видов наиболее перспективен в доминантном семействе чернотелок Tenebrionidae, которое представлено 9 видами, из которых наибольший интерес (по показателям удельного обилия, маршрутным учетам) представляют Anatolica gibbosa, Tentyria nomas, Blaps halophila.
Список литературы
- Хотько Э.И., Чумаков Л.С., Селявко Т.М. Функциональная структура населения жужелиц как показатель степени антропогенной нагрузки на экосистемы // Успехи энтомологии. — СПб.: Наука, 1993. — С. 72-74.
- Крыжановский О.Л. Состав и распространение энтомофаун земного шара. — М.: КМК, 2002. — 237 с.
- Ашихмина Т.Я. Биоиндикация и биотестирование — методы познания экологического состояния окружающей среды. — Киров: Наук. думка, 2005. — 246 с.
- Чигаркин А.В. Региональная геоэкология Казахстана. — Алматы: Қазақ ун-ті, — 224 с.
- ГиляровМ.С.MeTO/ibi почвенно-зоологических исследований. — М.: Наука, 1975. — 280 с.
- Кащеев В.А., Чильдебаев М.К., Псарев А.М. К методике изучения почвенной мезофауны членистоногих // Изв. МН-АН РК, 1997. — С. 30-37.
- Негробов С.О. Иллюстрированный определитель семейств жуков европейской части России. — Воронеж: Изд. ВГУ, 2005. — С. 103-104.
- МедведевГ.С. Определитель жуков-чернотелок Монголии // Тр. Зоол. ин-та АН СССР. — Л.: Наука, 1990. — 243 с.