В статье изучены особенности частоты встречаемости генов HLA II класса локусов DR, DQA1, DQB у больных ревматоидным артритом и ювенильным идиопатическим артритом в популяции русских Челябинской области. В указанных локусах авторами определены гены предрасположенности и устойчивости к развитию аутоиммунных ревматических заболеваний.
Заболеваемость аутоиммунными ревматическими заболеваниями и их распространенность существенно варьирует среди этнически и географически различных популяций во всем мире. Наиболее распространенным среди взрослого населения является ревматоидный артрит, а наиболее частым и самым инвалидизирующим ревматическим заболеванием у детей является ювенильный идиопатический артрит [1].
Ревматоидный артрит (РА) — аутоиммунное ревматическое заболевание неизвестной этиологии, характеризующееся хроническим эрозивным артритом и системным поражением внутренних органов [1]. Ювенильный идиопатический артрит (ЮИА) рассматривается как артрит неустановленной этиологии, присутствующий в течение 6 недель, возникший до 16-летнего возраста, при исключении других заболеваний [2]. От ревматоидного артрита взрослых ювенильный ревматоидный артрит отличается как по суставным, так и по внесуставным проявлениям [3].
РА и ЮИА соответствуют модели мультифакториального, полигенно-наследуемого заболевания, для которого являются характерными семейная подверженность, наличие наследственных маркеров предрасположенности к заболеванию, вариабельность клинических проявлений, зависящая от пола и возраста. К наследственным факторам, вовлеченным в процесс развития РА и ЮИА, традиционно относят генетическую предрасположенность, гормональный фон, расовую/этническую принадлежность, а к средовым — разнообразные внешние воздействия (вирусная и бактериальная инфекции, травма сустава, переохлаждение организма, инсоляция, лекарственные препараты и др.).
Роль наследственных факторов в развитии заболевания подтверждают различные иммуногенетические исследования [4, 5], при этом примерно 17 % для ЮИА и 40 % для РА от всей генетической составляющей принадлежит главному комплексу гистосовместимости человека — системе HLA [6, 7]. Детальный анализ ассоциаций РА и ЮИА с системой HLA демонстрирует, что HLA аллели ассоциируются не только с чувствительностью, но и с протекцией (устойчивостью) к развитию заболевания [8, 9].
Кроме этого, поиск генов предрасположенности, как и устойчивости к РА и ЮИА, проводимый во всем мире, привел к пониманию наличия межпопуляционных и межэтнических особенностей аллельного полиморфизма системы HLA, отражающих своеобразие условий проживания и образа жизни популяций в различных регионах мира [10]. Так, по данным многих авторов, заболеваемость и распространенность РА и ЮИА существенно варьирует среди этнически и географически различных популяций во всем мире. Известны регионы как с высокой (Англия, Швеция, Финляндия, Норвегия, Австралия, США), так и с низкой (Африка, Азиатские страны — Индия, Корея, Китай, Япония) частотой заболеваемости как для ЮИА, так и для РА. При этом даже в пределах одной страны заболеваемость РА и ЮИА отличается между различными этническими группами [5, 11].
Целью нашего исследования является анализ распределения генов HLA II класса локусов DRB1, DQA1, DQB1 у детей, страдающих ЮИА, и взрослых с диагнозом РА русской этнической группы Челябинской области.
Материал и методы
Группа больных ЮРА. Нами был обследован материал от 67 детей (67 % девочек и 33 % мальчиков) в возрасте 1-17 лет с установленным диагнозом ЮИА русской этнической группы. В соответствии с вариантом течения заболевания системный вариант отмечался у 22 пациентов (33 %); олигоартикулярный у 25 детей (37 %); полиартикулярный вариант верифицирован у 20 больных (30 %). Длительность заболевания к моменту исследования составила в среднем 4,8±4,3 года, манифестация первых клинических проявлений наблюдалась в среднем в 5,7±4,5 года.
Группа больных РА. Среди общего количества обследованных больных РА было 83 женщины (83 %) и 17 мужчин (17 %). Возраст больных был от 19 до 75 лет, средний возраст составил 51,5±12,0 лет. Возраст начала заболевания от 12 до 68 лет, средний возраст начала заболевания составил 40,7±13,1 года. Все пациенты были классифицированы по группам согласно рабочей классификации ревматоидного артрита.
В качестве контрольной группы использовали популяционную выборку, сформированную на основе ДНК-банка потенциальных доноров стволовой клетки ОГУП «Челябинская областная станция переливания крови». Контрольную группу составили 207 человек русской национальности, являющихся донорами Челябинской областной станции переливания крови и постоянно проживающих в Челябинской области. Одним из критериев отбора образцов было отсутствие родства между индивидуумами.
Типирование генов HLA I и II класса локусов DRB1, DQA1, DQB1 проводилось методом ПЦР с помощью наборов реагентов фирмы «ДНК-Технология» и набором сиквенс-специфических прай-меров [12].
Статистическая обработка включала расчет частоты встречаемости гена (Рх) и антигена (Ах%), рассчитываемая по закону Харди-Вайнберга, X2 критерий Пирсона с использованием поправки Йетса на непрерывность и критерий отношения шансов (OR).
Результаты и их обсуждение
Основой патогенеза ревматоидного артрита и идиопатического артрита является нарушение презентации антигенов. Доминантную роль в селекции и презентации антигенов в кооперации иммунных клеток играют гены HLA и кодированные ими антигены. Согласно литературным данным, в европеоидных популяциях установлена ассоциация ревматоидного артрита взрослых с генами локуса DRB1*01, *03, *04, а для ЮИА с генами B*27, DRB1*08, *11 [13-15].
В нашем исследовании при сравнении группы больных ЮИА нами определено статистически достоверное повышение частоты встречаемости гена DRB1*08 (р < 0,05, OR = 3,2, 95 % CI = 1,4-6,8). Сравнительный анализ распределения вариантов генов в локусе DQA1 показал, что в данной группе пациентов выявлено повышение частоты аллеля DQA1*04:01 (р < 0,05, OR = 3,3, 95 %CI = 1,4-7,5) относительно контроля. Результаты представлены в таблице.
В локусе DQB1 у детей с ЮИА повышена частота встречаемости аллеля DQB1*04:02 (р < 0,05, OR = 3,2, 95 %CI = 1,3-7,6). Результаты расчетов критерия отношения шансов (OR) и значение 95 % доверительного интервала свидетельствуют об ассоциативной роли этих генов в развитии ЮИА в популяции русских, проживающих на территории Челябинской области. При этом следует отметить, что наши данные согласуются с исследованиями, проведенными в различных европейских популяциях, например: Греции, Германии, Норвегии, Дании, Польше [10].
В группе больных РА установлено достоверное повышение частоты встречаемости гена HLA DRB1*04, чем в контрольной группе (57,00 % у больных РА против 18,81 % в контроле, р < 0,001, OR = 5,72), что определяет предрасполагающую роль этого гена в развитии РА. Эти данные совпадают с выводами работ о положительной ассоциации DRB1*04 c РА в большинстве европейских популяций [7, 16].
У больных РА, проживающих в Челябинской области, достоверно реже встречаются следующие варианты гена HLA DRB1: *11 (10 % против 18 %, X2 = 3,89, р < 0,05, OR = 0,48 при 95 %CI = 0,231,01), *13 (12,0 % против 27,7 %, X2 = 9,48, р < 0,05 OR = 0,36 при 95CI% = 0,18-0,70), *15 (13,0 % против 26,7 %, X2 = 7,31, р < 0,05, OR = 0,41 при 95 %CI = 0,21-0,80). Так как OR этих генов меньше 0,5 и эти значения с учетом 95 %-ного доверительного интервала меньше единицы, то полученные результаты позволяют рассматривать данные варианты HLA DRB1 как протекторные для развития РА у русских Челябинской области.
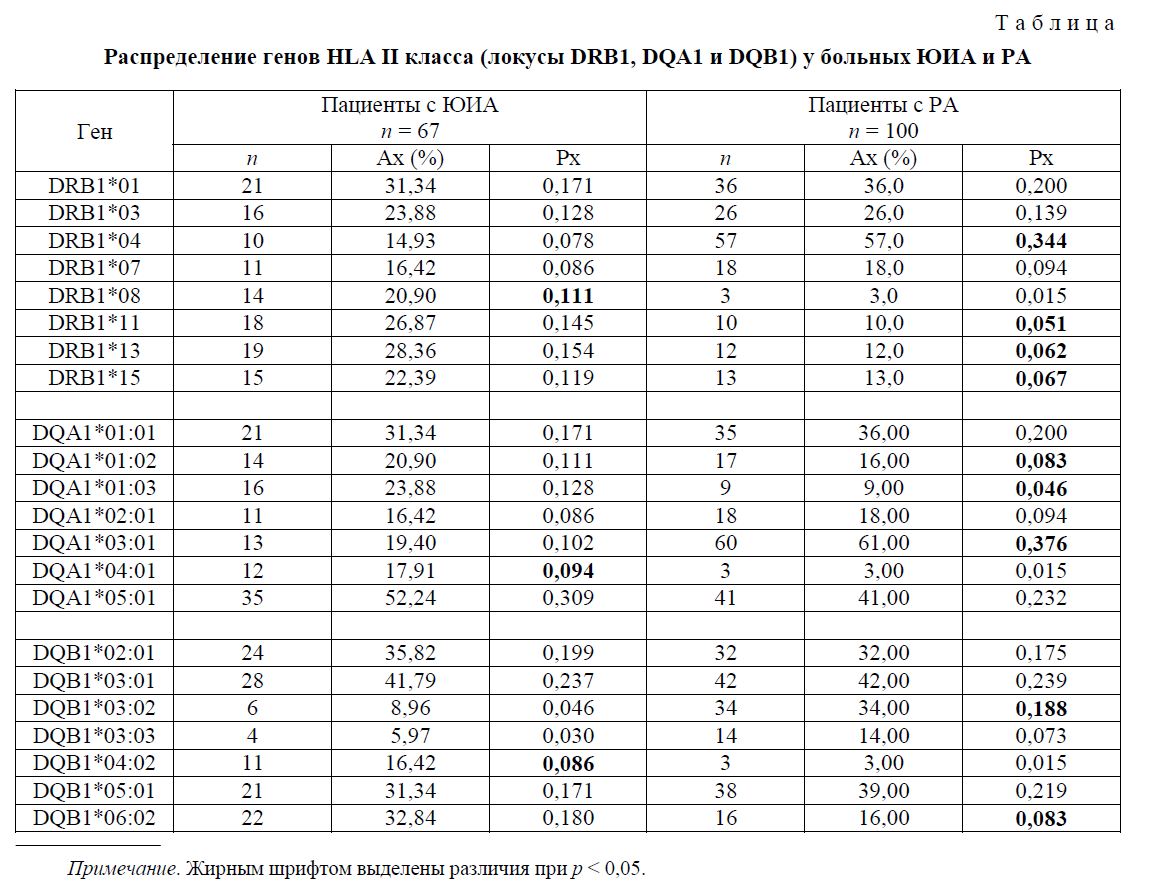
Сравнительный анализ распределения вариантов гена HLA DQA1 показал, что в группе больных РА достоверно повышена частота аллеля DQA1*03:01 относительно контроля (61,0 % против 23,3 %, X2 = 41,45, р < 0,001, OR = 5,16 95 %CI = 3,07-8,67). Аллели DQA1*01:02, *01:03 встречаются достоверно реже (x2 = 7,54, р < 0,05 и X2 = 5,36, р < 0,05), что позволяет предположить протективую роль этих аллелей в развитии предрасположенности к РА.
В локусе DQB1 у больных РА достоверно повышена частота аллеля *03:02 (34,0 % против 14,36 %, X2 = 15,63, р < 0,001, OR = 3,07, 95 %CI = 1,73-5,45). В меньшей степени оказалась повышенной частота другого аллеля DQB1*03:01. Однако это повышение не достигало уровня статистической значимости (р > 0,05). Достоверно и очень существенно снижена частота встречаемости аллеля DQB1*0602/8 в группе больных ревматоидным артритом (16,0 % против 40,1 %, X2 = 17,82, р < 0,001, OR = 0,28, 95 %CI = 0,16-0,52).
Полученные ассоциации определенных вариантов генов HLA различных локусов II класса с развитием ревматоидного артрита взрослых и ювенильного идиопатического артрита могут объясняться либо прямым включением HLA-антигенов в патогенез этих болезней (теория «молекулярной мимикрии», рецепторная теория, гипотеза общего эпитопа), либо существованием неравновесного сцепления между аллелями HLA и генами-регуляторами иммунного ответа (Ir-генами).
Точные механизмы ассоциации HLA-DRB1*08, DQA1*04:01, DQB1*04:02 с ЮИА еще не выяснены. По некоторым данным, HLA- DRB1*08 находится в неравновесном сцеплении с геном ТАР1В, который кодирует мембранные белки-переносчики, отвечающие за движение эндогенных антигенных пептидов по эндоплазматической сети и погрузку их на молекулы HLA I и II класса. Таким образом, абберации в ТАР-генах могут нарушать процесс доставки антигенных, в том числе и эндогенных, пептидов к молекулам HLA. Однако следует отметить, что именно молекулы HLA-DRB1 играют ведущую роль в изменении процесса презентации антигенов иммунокомпетентным клеткам. В результате происходит стимуляция выработки провоспалительных цитокинов Т-лимфоцитами, в том числе ФНО-а, которые, в свою очередь, стимулируют пролиферацию и дифференцировку В-лимфоцитов и продукцию аутоантител [4, 6].
Механизм ассоциации генов HLA-DRB1*04, DQA1*03:01, DQB1*03:02 с РА был предложен еще в 1987 г. П.Грегерсеном. Согласно его теории, варианты генов HLA-DRB1*04, которые ассоциированы с РА, кодируют синтез очень близких друг другу последовательностей аминокислот в положении 8-12 первой гипервариабельной области DRp1 цепи, образующей антигенсвязывающую бороздку. Эти последовательности получили название «общий эпитоп» (shared epitope — SE). Точные механизмы реализации «общего эпитопа» в развитии заболевания до конца не выяснены. Возможно, этот эпитоп с высокой степенью аффинности специфически связывает антигены синовиальных оболочек, что активирует иммунную реакцию аутореактивных клонов Т-лимфоцитов на собственные антигены суставной ткани, и таким образом предрасполагает к аутоиммунному артриту [17]. Но до сих пор не было идентифицировано такого пептида, который узнавался бы единственным Т-хелпером и выходил на однотипное развитие иммунного ответа в определенных органах-мишенях. Однако некоторые измененные варианты естественных пептидов приближаются к требуемой модели, делая это объяснение вполне вероятным. Однако такая же последовательность также присутствует на DRp 1 цепи HLA-DRВ1*03, но его роль в патогенезе РА в нашем исследовании не определена, что может объясняться этническими особенностями изучаемой популяции.
Помимо ассоциации с развитием аутоиммунных ревматических заболеваний в нашем исследовании определены протективные к развитию заболевания гены HLA: DRB1*11, DRB1*13, DRB1*15, DQA1*01:02, DQA1*01:03, DQB1*06:02. Подобные ассоциации были обнаружены в исследованиях других мировых популяциях — англичан, норвежцев, датчан [11]. Основываясь на этих наблюдениях, E.Zanelli сформулировал альтернативную гипотезу для механизма ассоциации между генами HLA II класса и РА — гипотеза резистентности РА, или RAP гипотеза (Rheumatoid АгЙіг^ Protective hypothesis) [18]. Согласно этой гипотезе, в молекуле DRB1, функция DERAA последовательности была не как части Р4 кармана, а как пептида, который может связаться с HLA-DQ молекулами и может быть узнан Т-регуляторными лимфоцитами (Treg+), которые подавляют индуцированные Т-клетки. Эта модель объясняет, почему DRB1*0103 и *0402-связанные гаплотипы не ассоциированы с РА, несмотря на носительство предрасполагающих DQ аллелей. Также DERAA-носительство DRp 1 цепей защищает против РА предрасположенности среди DQ5 позитивных индивидуумов и пересекается с прогрессированием заболевания среди DQ3 позитивных индивидуумов. Позднее D.Reviron классифицировал SE-негативные DRB1 аллели согласно электрическому заряду P4 кармана и исследовал ассоциацию между отрицательным или нейтральным зарядом c резистентностью к РА [19]. Другой подход был предпринят De Vries, который предложил новую форму SE модели, в которой наличие последовательности АА в положении 67-74 (вместо 70-74) прогнозирует или восприимчивость, или защиту в развитии РА [20]. Но на сегодняшний день ни одна из существующих гипотез не может полностью объяснить сложные отношения между генами HLA и РА или ЮИА.
Таким образом, полученные материалы позволяют предполагать, что определенные гены и аллели HLA II класса вовлечены в патологический процесс аутоиммунных ревматических заболеваний, но пути реализации этого процесса различны. Носительство предрасполагающего гена не является показателем неизбежности развития РА или ЮИА, но при соответствующем воздействии определенных факторов может выступать как одна из причин развития аутоиммунного процесса.
Список литературы
- НасоновЕ.Л. Ревматоидный артрит как общемедицинская проблема // Тер. архив. — 2004. — № 5. — С. 5-7.
- Зарецкая Ю.М., Леднев Ю.А.HLA 50 лет. — Тверь: Триада, 2008. — 152 с.
- Руководство по детской ревматологии / Под ред. Н.А.Геппе, Н.С.Подчерняевой, Г.А.Лыскиной. — М.: ГЭОТАР-Медиа, 2011. — 720 с.
- Glass D.N., Gianini E.H. JRA as a complex genetic trait // Arthritis. Rheum. — 1999. — 42. — Р. 2261-2268.
- TuressonС., Matterson EL. Genetic of rheumatoid arthritis // Mayo Clin. Proc. — 2006. — 81(1). — P. 64-101.
- Prahalad S., Glass D.N. A comprehensive review of the genetics of juvenile idiopathic arthritis // Pediatric Rheumatology. — 2008. — 6. — P. 11.
- Winchester R., Dwyer E., Rose S. The genetic basis of rheumatoid arthritis. The shared epitope hapothesis // Rheum. Dis. Clin. North Am. — 1992. — 18. — P. 761-783.
- Hollenbach J.A., Thompson S.D., Bugawan T.L., Ryan M., Sudman M., MarionМ., Langefeld C.D., Thomson G., Erlich H.A., Glass D.N. Juvenile Idiopathic Arthritis and HLA Class I and Class II Interactions and Age-at-Onset Effects // Arthritis & Rheuma — 2010. — Vol. 62, No. 6. — Р. 1781-1791.
- Van der Horst-Bruinsma I.E., Visser H., Hazes J.M., BreedveldF.C., Verduyn W., Schreuder G.M. HLA-DQ-associated predisposition to and dominant HLA-DR-associated protection against rheumatoid arthritis // Hum. Immunol. — 1999. — Vol. 60. — P. 152-158.
- Thomson W., Donn R. Genetic Epidemiology: Juvenile Idiopathic arthritis genetics — What's new? What's next? // Arthritis Res. — 2002. — 4. — P. 302-306.
- Shapira Y., Agmon-Levin N., Shoenfeld Y. Geoepidemioligy of autoimmune rheumatic diseases // Nat. Rev. Rheumatol. — 2010. — 6. — Р. 468-476.
- Downing J., Guttridge M.G., Thompson J., Darke C. Five-locus HLA typing of hematopoietic stem cell donor volunteers using PCR sequence specific primers // Genet. Testing. — 2004. — 8, No. 3. — P. 209.
- Morling N., Friis J., Fugger L., Geergsen J., Heilman C., Pedersen F.K., Bdum N., Svejgaard A. DNA polymorphism of HLA class II genes in pauciarticular juvenile rheumatoid arthritis // Tissue Antigens. — 1991. — Vol. 38, No. 2. — P. 16-23.
- Pascual M., Mataran L., Jones G. et al. HLA haplotypes and susceptibility to rheumatoid arthritis. More than class II genes // Scand J. Rheumatol. — 2003 — 32, No. 3. — P. 196.
- Smerdel A., Ploski R., Flato B. Juvenile idiopathic arthritis (JIA) is primarily associated with HLA-DR8, but not DQ4 on the DR8-DQ4 haplotype // Ann. Rheum. Dis. — 2002. — 61. — P. 354-357.
- Harney S., Wordsworth B.P. Genetic epidemiology of rheumatoid arthritis // Tissue Antige — 2002. — Vol. 60. —465-473.
- Gorman J., David-Vaudey E., Pai M. et al. Particular HLA-DRB1 shared epitope genotypes are strongly associated with rheumatoid vasculitis // Arthritis and rheumatism. — 2004. — 50, No. 11. — P. 3476-3484.
- Zanelli E., Gonzalez-Gay M.A., David C.S. Could HLA-DRB1 be the protective locus in rheumatoid arthritis // Immunol. Today. — 1995. — 16. — P. 274-278.
- Reviron D., Perdriger A., Toussirot E., Wendling D., Balandraud N., Guis S. Influence of shared epitope-negative HLA-DRB1 alleles on genetic susceptibility to rheumatoid arthritis // Arthritis Rheum. — 2001. — № — Р. 535-540.
- Vos K., Van der Horst-Bruinsma I.E., Hazes J.M.W., Breedveld F. C. et al. Evidence for a protective role of the human leukocyte antigen class II region in early rheumatoid arthritis // Rheumatology. — 2001. — № 40. — 133-139.