Изучены механизмы нейрогуморальной регуляции венозного кровообращения и лимфотока у рептилий. Исследованы афферентные влияния с барорецепторной зоны дуги аорты на венозное давление, венозный тонус и лимфодинамику у степных черепах. Показано участие венозных сосудов соматической и висцеральной отделов у рептилий в реакциях организма при действии нейрогенных факторов, что было выражено прессорными изменениями венозного давления и венозного тонуса. Результаты настоящего исследования направлены на развитие сравнительной физиологии кровообращения и лимфообращения.
Согласно многочисленным исследованиям, афферентные влияния с рецепторов дуги аорты приводят к рефлекторному изменению артериального давления, сосудистого тонуса и сердечной деятельности [1-7], а также лимфотока [8].
Относительно роли аортальной зоны и каротидного синуса в регуляции кровообращения и значения их в депрессорных реакциях у низших позвоночных мнения противоречивы.
При повышении давления в области каротидной зоны у жаб и лягушек наблюдались разряды в нервах каротидного синуса [9].
Некоторые авторы при повышении давления в каротидном синусе у жаб не наблюдали изменения в уровне кровяного давления и сердечной деятельности [10].
При повышении давления в изолированном участке бифуркации сонных артерий у черепах обнаружены как прессорные, так и депрессорные сдвиги артериального давления, а также увеличение лимфотока [11].
Отмечено, что у черепах в дуге аорты и в легочной артерии, а также в других артериях, иннерви-руемых нижними нервными стволами блуждающего нерва, локализованы баро- и хеморецепто-ры [12].
Существовало мнение, что депрессорный нерв низших позвоночных не является гомологом такового у млекопитающих [13].
У крокодилов была описана веточка от блуждающего нерва, идущая в дугу аорты и легочную артерию [14].
У ящериц и черепах также была обнаружена ветвь от блуждающего и верхнегортанного нервов, достигающая аорты и огибающая стенку желудочка сердца [9-12]. Этот нерв авторы считают аналогом депрессорного нерва у млекопитающих. Морфологически депрессорный нерв был идентифицирован и у других низших позвоночных: у щук [15], у лягушек [14], у змей [13].
Электрическая стимуляция гортанного нерва у жаб вызвала снижение или повышение артериального давления. Повышение давления в изолированном участке перфузируемого легочно-кожного ствола у жаб приводило к снижению артериального давления на 31 % с брадикардией или остановкой сердца [4-7]. У лягушек аналогичное раздражение гортанного нерва вызвало повышение давления в аорте и брадикардию [15].
Считают, что барорецепторы дуги аорты у рептилий (черепах), согласно результатам электронно-микроскопических исследований, сходны с таковыми у млекопитающих [16].
Стимуляция центральных концов стволовых нервов у черепах и ящериц приводила к падению артериального давления и частоты сердечных сокращений [15]. Автор заключил, что эти нервы сравнимы с депрессорным аортальным нервом млекопитающих. Позже другие авторы подтвердили этот факт [14].
Как видно из представленных данных, в литературе недостаточно сведений о развитии регуля-торных влияний афферентной зоны аортальной дуги у низших позвоночных на венозное кровообращение. Большим пробелом является недостаточность наших знаний о развитии нервной регуляции деятельности венозных сосудов у позвоночных, в частности, у низших позвоночных.
В настоящем разделе приводится материал, посвященный изучению роли барорецепторов дуги аорты на лимфоток и венозное кровообращение у представителей рептилий.
В серии опытов изучалось влияние раздражения барорецепторов дуги аорты на гемо- и лимфо-динамику у степных черепах. Проведено 28 опытов, включающих 101 наблюдение.
Результаты эксперимента показали, что при стимуляции барорецепторов дуги аорты (повышение давления до 60-80 мм рт ст.) у черепах наблюдалось повышение артериального давления в 37 % опытов от 20,08 ± 0,2 до 25,02 ± 0,6 мм рт ст., что составляло в среднем 125 % от исходного (р < 0,001), в 22 % артериальное давление снижалось, в среднем до 75 % от исходного фона (р < 0,001), в 41 % опытов сдвиги отсутствовали. Так же как и у лягушек, стимуляция барорецепторов дуги аорты у черепах вызывала угнетение сердечной деятельности. Однако брадикардия у черепах была менее выражена, чем у лягушек.
В венозных сосудах черепах отмечалось преимущественно повышение уровня давления. При этом чем дальше сосуд расположен от сердца, тем чаще наступали прессорные сдвиги венозного давления. Так, если в яремной вене давление повышалось в 46 % опытов, в подключичной — 55 %, брюшной и брыжеечной — в 60-68 % соответственно, то в бедренной вене в 80 % опытов сдвиги были прессорными. В остальных опытах давление в указанных венах либо не изменялось, либо понижалось (табл., рис. 1). Со стороны лимфатической системы у черепах наблюдались значительные сдвиги. Об этом свидетельствуют факты увеличения лимфотока в 65 % случаев на 20 % от исходного уровня и уменьшения его в 35 %. Сдвиги лимфотока наступали позже, чем кровяного давления и были более длительными (табл., рис. 1).
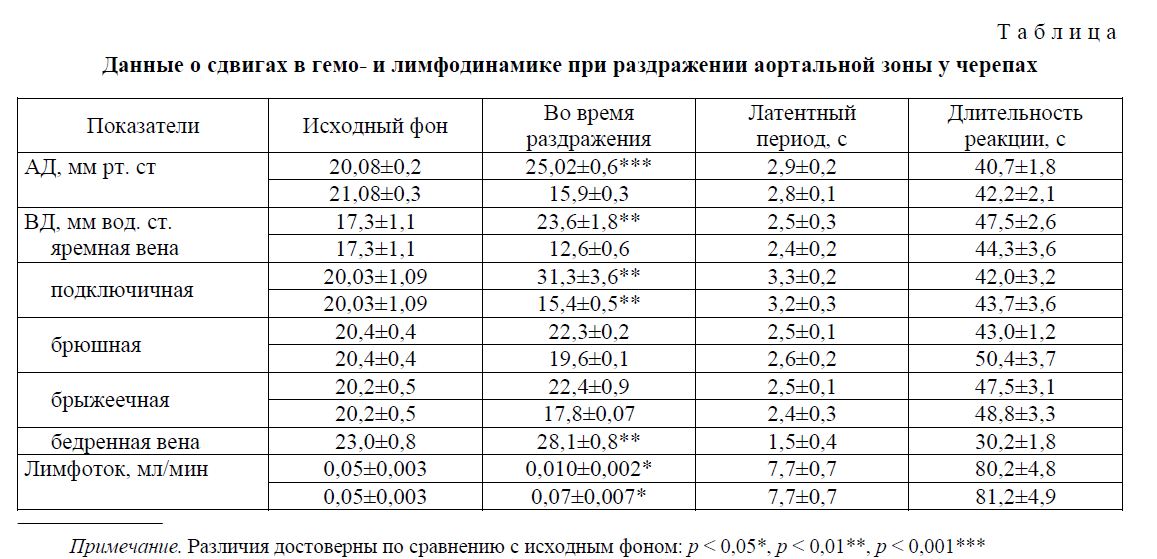
Данные о сдвигах артериального, венозного давления и лимфотока у черепах представлены в таблице. Давление в яремной вене повышалось на 6,3; в подключичной — на 11; в брюшной и брыжеечной — на 2 и в бедренной на 5 мм вод. ст. от исходного уровня. Наиболее выраженные сдвиги венозного давления наблюдались в яремной, подключичной и бедренной венах, т.е. в венах соматической области тела. В брюшной и брыжеечной венах величина повышения составляла 2-3 мм рт ст., т. е. сдвиги были слабыми. Сдвиги указанных показателей имели короткий латентный период и небольшую длительность, в пределах 1 мин.
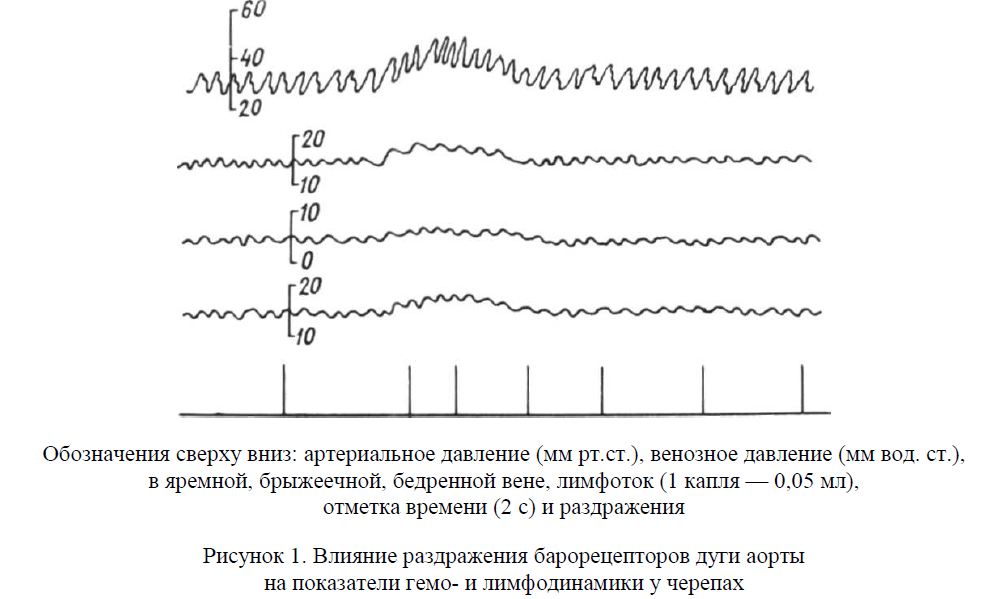
Рисунок 1. Влияние раздражения барорецепторов дуги аорты на показатели гемо- и лимфодинамики у черепах
На рисунке 1 видно, что при раздражении барорецепторов дуги аорты черепах наблюдалось повышение артериального давления от 17 до 43 мм рт. ст., венозного — в яремной от 15 до 20 мм вод. ст., брыжеечной — от 5 до 7 мм вод. ст., бедренной — от 16 до 19 мм вод. ст. и увеличение лим-фотока — в 3 раза от исходного фона.

На рисунке 2 представлен фрагмент опыта, проведенного на черепахе, где показано, что при стимуляции барорецепторов дуги аорты наблюдалось увеличение лимфотока на фоне депрессорной реакции артериального давления (от 34 до 12 мм рт. ст.) и повышения венозного давления (от 5 до 7 мм вод. ст.).
Как видно из приведенного выше материала, при раздражении барорецепторов дуги аорты черепах рефлекторно изменяется уровень артериального и венозного давления и величина лимфотока. У черепах при раздражении рецепторов дуги аорты артериальное давление повышалось в 37 % опытов, в 22 % оно снижалось, сопровождаясь брадикардией.
Таким образом, полученные данные свидетельствуют о прессорных влияниях с аортальной зоны низших позвоночных на кровяное давление, а также об отсутствии сформированной рефлексогенной депрессорной зоны в дугах аорт этих животных.
Как было указано выше, при стимуляции барорецепторов дуги аорты у черепах в большинстве наблюдений обнаружено повышение артериального и венозного давления, в небольшой части опытов — снижение артериального давления и увеличение лимфотока.
Наши данные подтверждают мнение [17], что аортальная зона рептилий не является гомологом депрессорной зоны дуги аорты у млекопитающих. Однако первые признаки депрессорных функций в этой зоне у черепах уже проявляются.
Как видно из полученных данных, при нейрогенных воздействиях наиболее выраженные сдвиги давления наблюдались в венах соматической области, а в венах брюшной области они были слабее. Данное явление можно объяснить тем, что в процессе филогенетического развития формирование венозных сосудов отстает от артерий. Согласно данным литературы, артерии и вены соматической области у низших позвоночных имеют сформированную адренергическую иннервацию, тогда как висцеральные вены и лимфатические сосуды еще содержат примитивные элементы адреналовой системы, а именно хромаффинные клетки, содержащие катехоламины [18, 19]. Эти видовые особенности иннервации венозных сосудов брюшной полости у низших позвоночных, а также слабый мышечный слой в стенках этих вен, вероятно, обусловливают более слабые реакции на нейрогенные воздействия, чем в венах шеи и конечностей.
References
- Euler U.S., von. Liljestrand G., Zotterman I. The excitation mechanism of the chemoreceptors of the carotid body // Scand. Arch. J. Physiol. — 1939. — 83. — № 1-3. — P. 132-152.
- Shapovalov L.M. About the features of hemodynamic changes caused by electrical stimulation of the depressor nerve in rabbits and cats // Physiol. Journal Ukr.SSR. — 1972. — 18. — № 1. — P. 65-69.
- Anyuhovsky E.P., Beloshapko G.G., Dremin S.A. Reactions of blood on the selective activation of myelinated and unmyelinated nerve fibers of the aortic // Bull. experts. biol. and med. — 1979. — 88. — № 11. — P. 515-517.
- Stinnett-Henry O., Peteson D.F., Bishop V.S. Rabbit cardiovascular responses to aortic nerve stimulation at reflexed carotid pressure // Amer. J. Physiol. — 1999. — 236. — № 5. — P. 769-774.
- Hartinkainen I., Ahonen E. et al. Effect of acute intravenous volume loading on haemodynamics and aortic baroreceptor activity in dogs // Acta Physiol. Scand. — 1989. — 135. — № 3. — P. 299-307.
- Timman I.A.K., Drinkhill M.I., Hainsworts R. Reflex to stimulation of mechanoreceptors in the left ventricle and coronary arteries in anaesthetized dogs // J. Physio — 1993. — Vol. 472. — P. 769-783.
- Varga K., Gantenberg N., Kunos G. Endogenous ү-aminobutyric caid (GABA) mediates ethanol inhibition of vagally mediated reflex bradycardia elicted from aortic baroreceptors // J. Pharmacol. and Exp. Ther. — 1994. — Vol. 268. — № 2. — P. 1057
- Musatova L.P. Changes in lymph flow, lymph protein composition and capacity of lymphatic vessels in acute vascular insufficiency // Mater. scient.-pract. conf. Inst. Physiol. AN KazSSR dedicated to the 100th anniversary of V.Lenin. — 1970. — P. 6265.
- Ishii K., Honda K., Ishii K. The function of carotid labyrinth in the toad // Tohoku J. Exp. Med. — 1966. — 88. — P. 103-106.
- T. Pressure and heart fate responses to raised carotid pressure in the toad // Amer. J. Physiol. — 1979. — Vol. 237. — № 6. — P. 639-643.
- Bulekbayeva L.E. et al. Cholinergic mechanisms of regulation of lymph and blood circulation in reptiles // DAN RK. Biological Series. — 1992. — № 1. — 73-76.
- Ishii K., Kusakabe T. Geoclemmys reevesii. Electrophysiological aspects of reflexogenic area in the chelonian // Respirat. Physiol. — 1985. — 59. — № 1. — P. 45-54.
- Mills T.W. The innervations of the heart of the slides terrapin (Pseudemys rugosa) // J. Physiol. — 1885. — 6. —246-286.
- Gaskell W.H. On the structure distribution and function of the nerves which innervate the visceral and vascular systems // J.Physiol. — London, — № 7. — P. 1-80.
- Fedele M. I nervi del tronco arteioso nel guardo della innervasione cardiac nei rettilt e il problema del «depressore» nee Ver-tebrata // Mem. R. Acc. Naz. Lincei. Ser. — 1937. — P. 387-520.
- Mills T.W. Some observations on the influence of the vagus and accelerators on the heart etc. of the tortoise // J. Physiol. — London, — № 5. — P. 359-361.
- I. The reptilian baroreceptor and its role in cardiovascular control // Am. J. Zool. — 1987. — № 27. — P. 111-120.
- Nicol J.A.C. Autonomic nervous system in lower chordates // Biol. Rev. — 1952. — 27. — P. 1-І9.
- SmittenSympatho-adrenal system in the phylo-ontogeny of vertebrates. — Moscow, 1972. — 346 p.