В статье приводятся данные, показывающие, что у экологически сходных животных конкуренция значительно ослабляется благодаря различиям во времени их активности (разделение активности днем и ночью). Показано также, что близкородственные виды животных характеризуются различными пищевыми потребностями. У некоторых симпатриантов одновременно обнаруживаются различия в использовании пространства, времени и пищи.
Полупустыня составляет 49% площади Азербайджанской Республики [1]. Именно здесь и сосредоточено ядро герпетофауны Азербайджана. Из 59 видов рептилий герпетофауны 44 вида представлены здесь [2]. Из этого количество видов 16 приходится на долю заурид (Sauria). Все они активные энтомофаги, большинство из них симпатричны и в их рационе присутствуют одни и те же компоненты [3]. Анализируя пищевой рацион некоторых симпатричных видов можно выявить степень трофической конкуренции между ними.
МАТЕРИАЛ И МЕТОДИКА
Из 9 исследованных видов были сделаны выборки около 100 особей: каспийский голопалый геккон - Cyrtopodion caspius (Eichwald, 1831), кавказская горная агама - Laudakia caucasica Eichwald, 1831, желтопузик - Pseudopus apodus Pallas, 1775, длинноногий сцинк – Eumeces schneideri (Daudin, 1802), быстрая ящурка - Eremias velox (Pallas, 1771), разноцветная ящурка - Eremias arguta Pallas, 1773, азербайджанская скальная ящерица - Darevskia raddei (Boettger, 1892), змееголовка стройная - Ophisops elegans Menetries, 1832, полосатая ящерица - Lacerta strigata Eichwald, 1831.
Пищевые ингредиенты устанавливались путем изучения непереваренных остатков в фекалиях животных по Вержуцкому, Журавлеву [4]; Целлариусу [5], а также изучением содержимых желудков коллекционных материалов. Определение проводилось по определителю насекомых [5].
Интенсивность конкуренции за пищевые ресурсы определялась по индексу Мориста [7]:

РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Современная герпетофауна республики включает 59 видов относящихся к 33 родам, 12 семействам и 3 отрядам. В аридных и семиаридных ландшафтах встречается 44 (75%) вида рептилий. По характеру питания пресмыкающиеся делятся на фитофагов, энтомофагов, ихтиофагов, батрахофагов, герпетофагов, орнитофагов и териофагов. В приведенной ниже таблице 1 выделен характер питания для видов рептилий аридных и семиаридных ландшафтов.
Таблица 1 - Особенности питания рептилий аридных и семиаридных ландшафтов Азербайджана
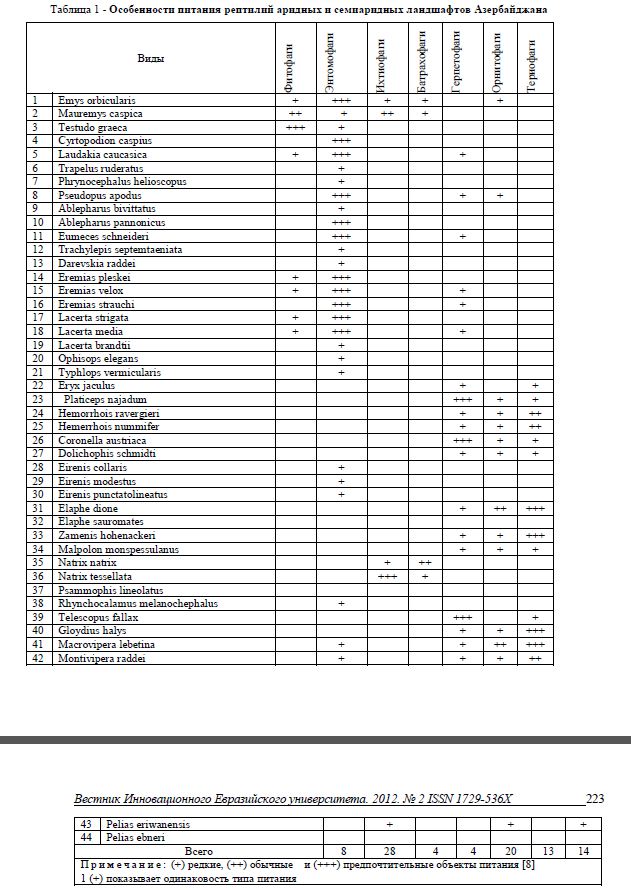
Среди этих видов только средиземноморская черепаха (Testudo graeca) является растительноядной. В пищевом рационе каспийской черепахи (Mauremys caspica) и болотной черепахи (Emys orbicularis) растения, особенно высшие водные растения, также играют определенную роль. Животная пища составляет 90% корма болотной черепахи. Это в основном водяные насекомые и особенно их личинки, а у каспийской черепахи основную долю составляют растения (80%) и небольшую часть - рыбы (20%). В наземной среде основную часть корма каспийской черепахи составляют прямокрылые, но в том же биотопе болотная черепаха отдает предпочтение паукообразным [9]. Различия в пищевом рационе видов, обитающих в одном биотопе, снижает конкуренцию между видами, что было подтверждено нами на территории около поселка Пирекюшкюль (Гобустан).
Большинство змей - эврифаги, в рационе которых встречаются все амниоты. К ним относятся Hemorrhois nummifer, H. ravergieri, Dolichophis schmidti, Elaphe dione, Macrovipera lebetina obtusa, Montivipera raddei и т.д. Следующие виды, такие как Zamenis hohenackeri, Z. persicus, Malpolon monspessulanus, отдают предпочтение грызунам и птицам, а Platyceps najadum, Coronella austriaca, Psammofis lineolatus, Telescopus fallax - грызунам [8].
Виды рода Natrix характеризуются как виды ихтиофаги, батрахофаги. Обыкновенный уж отдает предпочтение лягушкам, тогда как водяной уж в основном питается рыбами. Это также ослабляет конкуренцию между ними в одинаковой среде [10], что и было нами отмечено в некоторых водоемах.
Несмотря на наличие в корме змей и грызунов, такие виды, как Platyseps najadum, Coronella austriaca, Malpolon monspessulanus, являются заурофагами [8].
В исследуемом комплексе у 28 видов (64%) значительную долю в питании составляют беспозвоночные, основная доля (92%) которых приходится на наземных беспозвоночных.
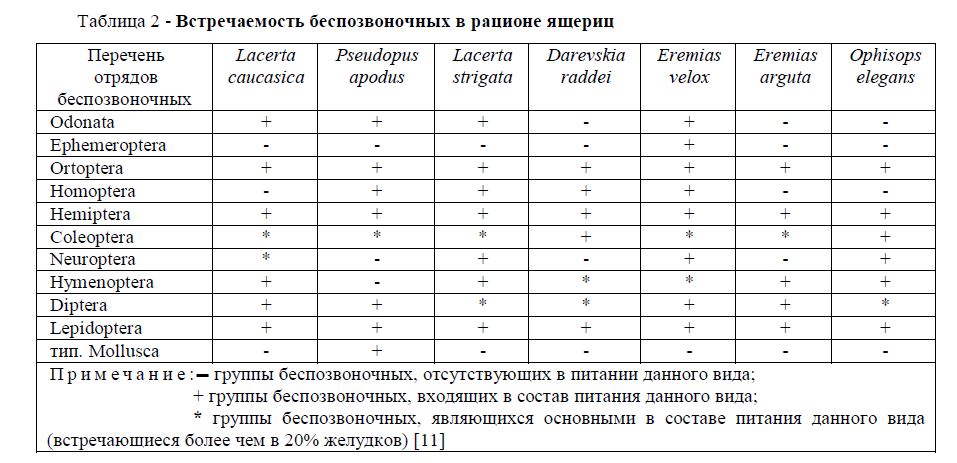
В результате анализа встречаемости беспозвоночных в рационе ящериц было выявлено, что предпочтение отдается следующим видам – это группы Ortoptera, Hemiptera, Lepidoptera, которые встречаются у всех рассмотренных видов пресмыкающихся. Как видно из таблицы 2 для всех изученных видов ящериц основными видами являются представители отряда Coleoptera. Самый богатый рацион корма обнаружен у быстрой ящурки. Наименьшая встречаемость относится к представителям отряда Ephemeroptera и типа Mollusca. Первый из них, хотя и незначительно, входит в состав пищевого рациона только быстрой ящурки. Представители типа Mollusca входят в кормовой рацион только желтопузика. Причиной этого является склонность желтопузика к более влажным биотопам. Наименьшее количество по разнообразию отрядов беспозвоночных выявлено в пищевом рационе разноцветной ящурки. Возможно, это объясняется слабой подвижностью данного вида, по сравнению с остальными рассмотренными ящерицами.
Таблица 2 - Встречаемость беспозвоночных в рационе ящериц
У некоторых симпатрических видов одного рода мы обнаружили различия в использовании пространства, времени и пищи. Во многих биотопах быстрая ящурка и разноцветная ящурка обитают совместно. Анализ литературных данных [9] и коллекционных материалов Института Зоологии НАН Азербайджана показал, что во многих исследованных пунктах на Апшероне они обитают совместно (в окрестностях поселков Зых, Говсаны, Пирекюшкюль и др). Однако во время экспедиций в указанные районы мы повсеместно встречали быструю ящурку, а разноцветная ящурка нами не была зарегистрирована. Лишь в Ширванском Национальном парке за однодневную экскурсию насчитали 25 быстрых ящурок и только 1 разноцветную.
Пищевой рацион быстрой и разноцветной ящурок был изучен на основании собственных сборов и материалов Института Зоологии АН Азербайджана. Подсчеты показали, что индекс сходства пищевого рациона обоих видов низок (Jλ =0,33 (рисунок 1)). Это объясняется тем, что они занимают различные микростации, несколько сдвинутым временем суточной активности, а также несколько различным пищедобывательным поведением.
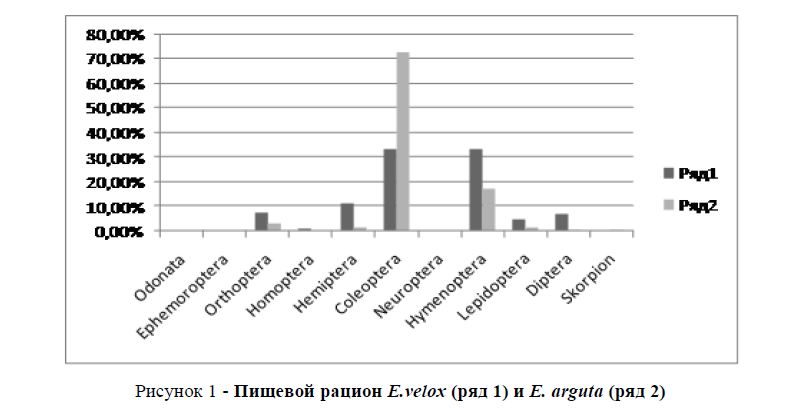
Рисунок 1 - Пищевой рацион E.velox (ряд 1) и E. arguta (ряд 2)
Таким образом, можно констатировать, что конкурентные взаимоотношения не являются причиной низкой численности разноцветной ящурки. Считаем, что разноцветная ящурка не адаптировалась к изменению экологических условий в местах обитания обоих видов в последние годы. На численности более пластичной быстрой ящурки эти изменения пока не отразились.
Кавказская горная агама распространена в республике интразонально от высот 300 м до высот 2500 м над уровнем моря. В Апшеронском и Гобустанском районах она симпатрична каспийскому голопалому геккону. Однако в отличие от кавказской агамы для каспийского голопалого геккона характерна ночная активность, что и устраняет возможность конкуренции. Характерными местообитаниями каспийского геккона являются расщелины скал, щели каменных заборов, стены различных строений, затененные места. Этот вид среди ящериц Апшерона является единственным видом, ведущим ночной образ жизни. Следовательно, кормовыми объектами являются активные ночные беспозвоночные, как то: саранчовые, долгоносики, жужелицы, сверчки, клопы и пауки.
В тех биотопах, где совместно обитают горная агама и быстрая ящурка, численность последней низка. Мы наблюдали такую ситуацию в Гобустанском заповеднике и на г. Гаргабазары. Индекс сходства пищевого рациона высок (Jλ =0,75). Возможно, это объясняется тем, что основным кормом для обоих видов являются представители отряда Coleoptera (рисунок 2). Например, в пищевом рационе у агамы они составляют 36,7%, у быстрой ящурки – 33,8%. На низкую численность ящурки, возможно, влияет то обстоятельство, что молодью ящурок питается кавказская агама.
Во многих горных и предгорных районах республики кавказская агама обитает совместно со скальной ящерицей и змееголовкой стройной. Это мы наблюдали и во время наших посещений плато Зуванд на юго-востоке республики. Здесь они встречаются на высотах до 1200-1800 м над уровнем моря.
Кавказская агама и скальная ящерица обитают в сходных микростациях. Однако индекс сходства в составе кормов очень низок (Jλ = 0,28). В пищевом рационе кавказской агамы преобладают представители отрядов Coleoptera (36,7%), Neuroptera (22%); а основным кормовым объектом азербайджанской ящерицы являются Hymenoptera (32,3%) и Diptera (18%) (рисунок 3). Кавказская агама охотится, в основном на скалах, редко спускаясь к подножию, а азербайджанская ящерица обычно у подножия и в травянистом покрове. Следовательно, в данном случае конкуренция за ресурсы либо ослабляется, либо устраняется.
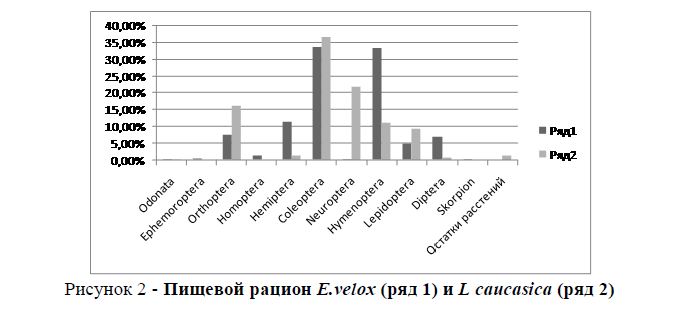
Рисунок 2 - Пищевой рацион E.velox (ряд 1) и L caucasica (ряд 2)
В случае перекрывания пищевых ниш обсуждаемых видов между ними возникают отношения хищник- жертва. Как отмечает И.С. Даревский, на Южном Кавказе сеголетки и молодые особи азербайджанской ящерицы становятся добычей кавказской агамы [12].
В Зуванде стройная змееголовка обитает на невысоких холмах каменистых участков, а также среди зарослей полыни. Ее можно встретить также среди зерновых и травянистых участков. Индекс сходства состава кормов кавказской агамы и змееголовки - Jλ = 0,46. Такой низкий индекс сходства объясняется различными микростациями в одинаковых биотопах.
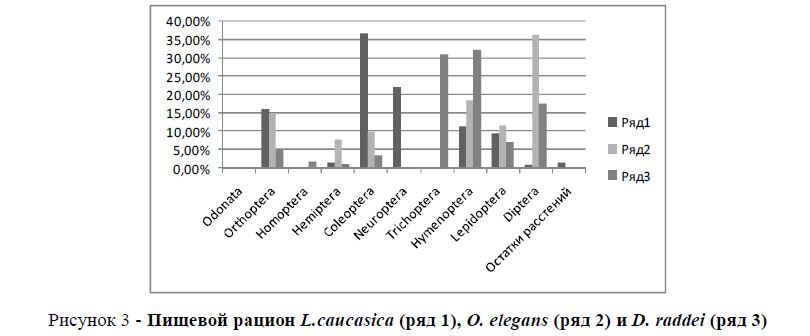
Рисунок 3 - Пищевой рацион L.caucasica (ряд 1), O. elegans (ряд 2) и D. raddei (ряд 3)
Индекс сходства в составе кормов стройной змееголовки и азербайджанской ящерицы очень высок - Jλ = 0,63. Объясняется это сходством первостепенных и второстепенных объектов питания. Первостепенным кормом змееголовки являются представители отряда Diptera – 36,4%. Последние в рационе азербайджанской ящерицы, имея второстепенное значение, составляют 18%. Основным кормом азербайджанской ящерицы являются представители отряда Hymenoptera (32,3%). Эта группа беспозвоночных является второстепенным кормом для змееголовки и составляет 18,6%.
Симпатричными видами являются желтопузик и полосатая ящерица. У обоих видов в кормах первостепенное значение имеют представители отряда Coleoptera: у желтопузика – 52,7%, у полосатой ящерицы – 35,6% (рисунок 4). Индекс сходства этих видов кормов составляет Jλ = 0,82
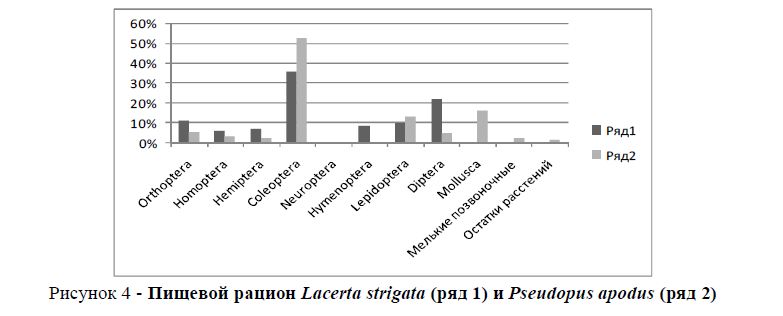
Рисунок 4 - Пищевой рацион Lacerta strigata (ряд 1) и Pseudopus apodus (ряд 2)
Желтопузик и длинноногий сцинк - крупные, подвижные ящерицы. Встречаются в степных и полупустынных биотопах, в зоне предгорий с каменистыми россыпями и со скудной растительностью. Совместно с ними обитают змееголовка стройная, быстрая и разноцветная ящурки. В желудках желтопузика и длинноногого сцинка довольно часто встречаются змееголовки и ящурки.
Внутри популяции также существуют различные адаптации, устраняющие или ослабляющие конкурентные отношения между особями. Например, в популяции змееголовки стройной, обитающей в Зуванде, летом особи становятся активными к 8-830 при температуре 19-200С. Первыми после ночевки появляются молодые особи. Взрослые особи появляются к 11 часам при температуре 23-240С.
Литература
- Керимов Н.К. Физико-географическое (ландшафтное) районирование Азербайджанской ССР / Тез.докл.научн.сессии, посв. 36-й годовщине установл. Советской власти в Азербайджане. - Баку,– С.11-13.
- Аскерова С.А. Особенности распространения рептилий полупустынных ландшафтов Азербайджана // Материалы XV республиканской научной конференции докторантов и молодых ученых. - Баку,– С. 64-65.
- Джафарова С.К. Структура биотопических, временных и пищевых гильдий фауны ящериц Азербайджана // Вестник Бакинского Университета. Серия естественных наук. - – № 2. – С. 86-91.
- Вержуцкий Б.Н., Журавлев В.Е. Щадящий метод изучения трофического спектра рептилий // Вопросы герпетологии. –1977. – С. 58-59.
- Целлариус А.Ю. Изучение питания ящериц по экскрементам // Вопросы герпетологии. – 1977. – 220 с.
- Определитель насекомых Европейской части СССР. - Л.: Наука, – 510 с.
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. - Москва: Наука, – 287 с.
- Туниев Б.С., Орлов Н.Л., Ананьева Н.Б., Агасян А.Л. Змеи Кавказа: таксономическое разнообразие, распространение, охрана. - М.: Товарищество научных изданий КМК, 2009. – 223 с.
- Алекперов А.М. Земноводные и пресмыкающиеся Азербайджана. - Баку: Элм, 1978. – 264 с.
- Fillippi & Capula M. & Luisseli L. & Agrimi U. The prey spectrum of Natrix natrix Linnaeus, 1758) and Natrix tessellata (Laurenti, 1768) in sympatric populations (Squamata: Serpentes: Colubridae) // Herpetozoa 8 (3/4). - С. 155-164.
- Бутов Г.С., Хицова Л.Н. Особенности питания земноводных и пресмыкающихся в антропогенных биотопах г. Воронежа // Вестник ВГУ. Серия Химия, Биология, Фармация. - – № 2. – С. 108-115.
- Даревский И.С. Скальные ящерицы Кавказа. - Ленинград: Наука,1967. – 213 с.