Листовая или бурая ржавчина является самой распространенной болезнью мягкой пшеницы в Казахстане. Возбудитель болезни – облигатный гриб Puccinia triticiana Eriks. (синоним –Puccinia recondita Rob.ex Desm. f.sp. tritici.) [1]. Одной из причин массового развития этой болезни является возделывание сортов пшеницы однотипных по устойчивости и появление новых вирулентных патотипов гриба, что неизбежно приводит к быстрой потере их иммунитета. Известно, что выявление путей снижения вредоносности данного гриба является создание ржавчиноустойчивых сортов пшеницы, и при этом эффективная значимая защита от листовой ржавчины может достигаться только при наличии и на основе достаточно полных сведений и знаний о популяции патогена [2, 3].
По литературным данным у возбудителя листовой ржавчины пшеницы насчитываются более 200 рас (патотипов) [4], которые отличаются своей агрессивностью и вирулентностью к определенным сортам. Представленность того или иного изолята на сорте зависит от взаимодействия генотипа патогена с генотипом растения-хозяина. В этой связи возникла необходимость изучение патогенных свойств возбудителя P.triticiana на сортах пшеницы.
Изучение стуктуры популяции листовой ржавчины из различных регионов Казахстана показало, что патоген отличается значительным генетическим разнообразием и высокой вирулентностью. Проанализировано 230 монопустульных изолятов гриба, из них идентифицировано 30 патотипов, имеющих от 5 до 13 генов вирулентности. В ходе анализа нами не выявлено изолята, вирулентного к гену Lr25, что показывает на высокую эффективность. Также можно отметить эффективные гены Lr9 и Lr19 у которых число изолятов способных преодолеть устойчивость, обусловленную этими генами, было низким. Так, коэффициенты вариации частот встречаемости изолятов, вирулентных к указанным генам составили 13,3% и 23,3%, соответственно. Коэффициенты вариации частот встречаемости изолятов, вирулентных к остальным генам устойчивости колебались от 40,0% до 83,3% (таблица 1).
Таблица 1 Характеристика Lr-линии по устойчивости к изолятам листовой ржавчины пшеницы в Казахстане
|
Lr-линии |
Частота встречаемости изолятов возбудителя листовой ржавчины |
|||
|
Вирулентные, шт. |
% |
Авирулентные, шт. |
% |
|
|
|
|
|
|
|
|
Lr1 |
25 |
83,3 |
5 |
16,7 |
|
Lr2a |
24 |
80,0 |
6 |
20,0 |
|
Lr2c |
24 |
80,0 |
6 |
20,0 |
|
Lr3 |
20 |
66,7 |
10 |
33,3 |
|
Lr3ka |
22 |
73,3 |
8 |
26,7 |
|
Lr9 |
4 |
13,3 |
26 |
86,7 |
|
Lr11 |
6 |
20,0 |
24 |
80,0 |
|
Lr16 |
23 |
76,7 |
7 |
23,3 |
|
Lr17 |
25 |
83,3 |
5 |
16,7 |
|
Lr19 |
7 |
23,3 |
23 |
76,7 |
|
Lr20 |
18 |
63,3 |
12 |
40,0 |
|
Lr24 |
12 |
40,0 |
18 |
60,0 |
|
Lr25 |
0 |
0 |
30 |
100,0 |
|
Lr26 |
20 |
66,7 |
10 |
96,7 |
|
Lr29 |
21 |
70,0 |
9 |
30,0 |
|
Lr30 |
23 |
76,7 |
7 |
23,3 |
|
Среднее |
17,2 |
57,3 |
12,8 |
42,7 |
Высокой частотой встречаемости в популяции характеризовались гены вирулентности Lr1, Lr2а, Lr2с, Lr3ка, Lr16, Lr29 (70,0-83,3%), средней концентрацией (40,0-66,7%) отличились гены Lr3, Lr20, Lr24 и Lr26. Изучение влияния генотипа сортов пшеницы на структуру популяции патогенов показало, что каждый из сортов имеет определенное специфическое давление на популяцию возбудителя. Так, с сорта Саратовская 29 выявлены патотипы имеющие разные вирулентные способности (THTK-68,7%; THPH-60,0%; SFPH-55,0%; THPC-65,0%). Приуроченность патотипов листовой ржавчины к сорту Саратовская 29 может оказать существенное влияние на развитие эпифитотии, особенно в тех регионах республики, где посевами этого сорта заняты значительные площади. Такая же тенденция наблюдается и на сорте Стекловидная 24, которая считается одним из высокоурожайных сортов на юге Казахстана, где большая часть посевов доминирует в структуре зерновых культур. Следовательно, с увеличением площадей под этим сортом возможно накопление и дальнейшее распространение патотипов TFPH, SBPC и SFHH.
Кроме того, на сортовых уредообразцах найдены патотипы TKPH (Реке, Нуреке, Э-17) и TTPH (Караспан, Тритикале Т-10), что свидетельствует об их специализации к песпективным сортам пшеницы и тритикале, а также не исключена возможность их развития и распространения на коммерческих сортах пшеницы. Патотипы SHKM (62,5%), HGKB (37,5%) и FQHQ (56,3 %) способствовали поражению тритикале листовой ржавчиной на юго-востоке республики с характерными особенностями к изогенным Lr-линиям, хотя по литературным данным пшенично-ржанной гибрид (тритикале) в отношении к листовой ржавчине показывает устойчивость. Однако в последние годы данная культура проявляет высокую чувствительность к патогену. Так, выделенные патотипы SHKM (Lr1, 2а, 2с, 16, 26, 11, 17, 30, 19, 29) и FQHQ (Lr2а, 2с, 3, 9, 3ка, 17, 30, 19, 29) обладают универсальной вирулентностью, генотипы этих патогенов содержат высокую концентрацию известных генов вирулентности, кроме гена Lr25. Патотип HGKB (Lr2а, 3, 16, 11, 17, 30) показывающий на стандартном наборе лишь отдельные восприимчивые гены устойчивости, были авируленты к дополнительному набору Lr19, 20, 25 и 29 являющий дифференцирующий способностью для условий Казахстана. Кроме того, в северном Казахстане широкое распространение имеет сорняк лещица (Isopyrum fumaroides), который считается промежуточным хозяином данного гриба и он может играть некоторую роль в возобновлении и изменчивости популяций. Патотип TRP/G, выделенный из указанного растения, отличается по вирулентности к изогенным линиям листовой ржавчины. Патотипы PHTB (Lr1, Lr2c, Lr3, Lr16, Lr26, Lr3ka, Lr11, Lr17, Lr30) и KHPF (Lr1, Lr2a, Lr2c, Lr3, Lr16, Lr26, Lr3ka, Lr11, Lr17, Lr30, Lr25, Lr29) входящие в состав дербенской популяции, отличаются высокой вирулентностью к Lr-линиям. Особенно, можно отметить второй патоген, который проявил вирулентность к эффективному гену Lr25.
Таким образом, проведенный нами в последние годы мониторинг популяции гриба по признаку вирулентности, показывает значительные изменения вирулентности идентифицируемых изолятов. Простые патотипы с 1 или 2 генами вирулентности в популяции полностью отсутствуют, в основном популяцию гриба составляют патотипы с 5 и более генами. Гены вирулентности Lr1, Lr2a, Lr2c, Lr3, Lr16, Lr3ka, Lr17, Lr30, Lr29 встречаются на разных изолятах выделенных как из коллекционных, так и из производственных посевов. В природной популяции определились сложные патотипы с 8-13 генами вирулентности, и они отмечены как в годы эпифитотийного, так и депрессивного развития болезни. Следовательно, их распространению в природе способствует их генетическая приспособленность к сортам пшеницы и другим растениям, выраженная высокой вирулентной способностью патотипов. Редко встречающие патотипы как правило, имеют низкую вирулентную способность и ее могут снизить более доминирующие патотипы в популяции, которые имеют высокие вирулентные свойства.
Неспецифическая устойчивость считается количественным признаком, и она проявляется в уменьшении репродуктивной способности патогена, в уменьшении числа пустул на единицу листовой поверхности и увеличении длительности латентного периода. Эти показатели определяют агрессивность патогена и в лабораторных условиях нами были изучены данные параметры с целью выявления конкурентоспособности полученных патотипов из природной популяции на сортах яровой и озимой пшеницы. Результаты опытов показали, что продолжительность латентного периода развития листовой ржавчины на растениях в фазе проростков составила 9-12 суток. Малым промежутком времени проявление болезни на определенном участке листа наблюдалось при заражении патогеном KHPF, среднее значение составило 9,1 сут (таблица 2).
Таблица 2 Изучение латентного периода патотипов возбудителя P.triticiana на сортах пшеницы
|
Название сорта |
Латентный период (сутки) |
||||
|
THTK |
PHTB |
KHPF |
TRPG |
FKLM |
|
|
Акмола 2 |
10 |
12 |
9 |
9 |
9 |
|
Байтерек |
9 |
10 |
9 |
9 |
9 |
|
Омская 29 |
9 |
10 |
9 |
9 |
10 |
|
Алтайская 100 |
10 |
12 |
9 |
10 |
9 |
|
Э- 791 |
10 |
11 |
9 |
10 |
10 |
|
Э -809 |
11 |
12 |
9 |
10 |
9 |
|
Саратовская 29 |
11 |
11 |
9 |
10 |
11 |
|
Алмалы |
10 |
11 |
10 |
10 |
9 |
|
Стекловидная 24 |
10 |
11 |
9 |
10 |
10 |
|
Среднее |
10,0 |
11,1 |
9,1 |
9,7 |
9,6 |
Медленное развитие болезни на проростках характеризуется для патотипа PHTB, где отдельные сорта (Акмола 2, Алтайская 100 и Э-809) имели более длинный период – 12 суток. На остальных образцах развитие патогена гриба (THTK, TRPG и FKLM) при ежедневном подсчете уредопустул отмечается незначительная разница в промежутке времени, т.е. 10,0, 9,6 и 9,7 суток, соответственно.
Замедленное развитие патогена PHTB на изучаемых сортах пшеницы привело к уменьшению числа его поколений, и спороношение гриба было слабым, при этом выход спор с листовой поверхности было минимальным 0,003-0,020 г. В этом случае патоген PHTB считается слабо агрессивным, так как не обладает определенным комплексом свойств агрессивности. Аналогичные результаты получены при изучении патотипа THTK и поэтому его также можно отнести к слабо агрессивному патогену. Хотя оба патогена считаются вирулентными. Патогенность и вирулентность это постоянные, неподверженные колебаниям свойства возбудителей болезни. Уровень же агрессивности одного и того же патогена может изменяться в зависимости от условий развития и других причин. Таким образом, результаты опыта показали, что не всегда вирулентные патотипы обладают агрессивностью. Признак агрессивности проявляется в патотипах со средней вирулентностью KHPF, TRPG и FKLM, поскольку они обладают определенными патогенными свойствами: высокой энергией размножения (споруляцией), способностью заражать растения минимальным количеством инфекционного начала, малой продолжительностью процесса заражения и инкубационным периодом. При этом инкубационный период болезни на растениях составил 9 суток, при среднем показателе инфекции 3,2-3,4 балла, в отдельных случаях на отдельных сортах их реакция имела 3,6-3,8 балла. Следовательно, репродуктивная способность у них была высокой (0,034-0,042 г.) по сравнению с патотипами THTK и PHTB (0,007-0,017 г.) (Рис1).
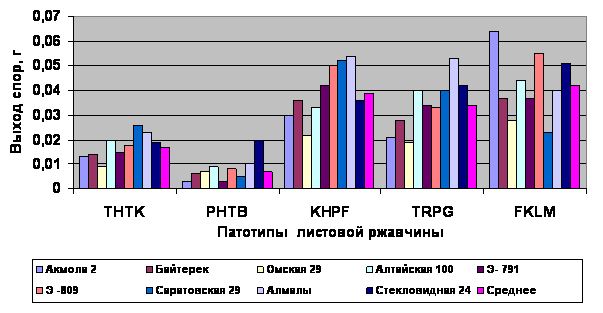
Рис. 1. Выход спор гриба с листовой поверхности на сортах пшеницы
Таким образом, патогены гриба KHPF, TRPG и FKLM можно определить как высоко агрессивные патотипы и присутствие их в природной популяции при благоприятных погодных условиях может вызвать высокую степень развития болезни, создать эпифитотию гриба на посевах пшеницы.
Кроме того, агрессивность гриба по отношению к сортам пшеницы можно измерить таким показателем как эффективность клонирования (ЭК). Ее определяют путем сравнения различных изолятов гриба по количеству пустул, проявившихся на единице листовой поверхности, нанеся на нее дозированное количество спор гриба [5]. Изучение данного показателя показало, что ЭК на одном и том же сорте независимо от его устойчивости сильно варьировала в зависимости от используемого патотипа гриба. Например, показатели ЭК патотипов THTK, PHTB, KHPF, TRPG и FKLM на проростках сорта Акмола 2 имели достоверные отличия в процентном соотношении 1,55; 0,42; 2,48; 1,93; 2,58 соответственно (Рис.2).
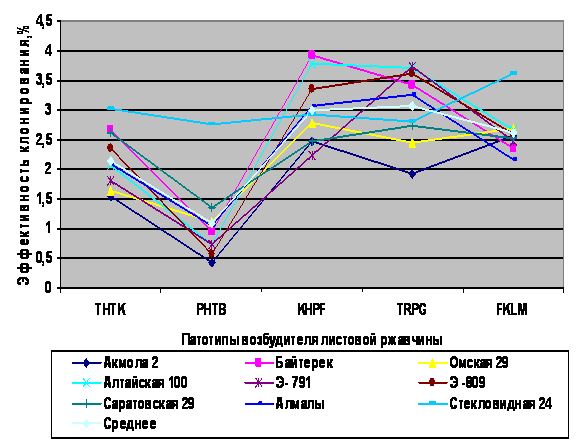
Рис. 2. Изучение свойств агрессивности листовой ржавчины на сортах пшеницы
с помощью показателя эффективности клонирования
Аналогичные данные по признаку имели и другие исследуемые сорта, следовательно, первые два патотипа являются слабоагрессивными, а три другие – высоко агрессивными, что подтверждает эффективность клонирования. При этом высоко агрессивные патотипы способны не только заразить растение малым количеством спор, но и быстро осуществить заражение, при коротком инкубационном периоде, образовав в течение лета несколько генерации бесполых спороношений, которые легко распространяются ветром и служат для массовых повторных заражений.
Таким образом, при изучении патогенных свойств возбудителя P.triticiana по признаку вирулентности было выявлено, что популяция листовой ржавчины пшеницы в Казахстане состоит из патотипов с 8-13 генами вирулентности. Встречаемость изолятов к эффективным генам Lr9 и Lr19 были низкими, не обнаружено ни одного изолята к эффективному гену Lr25. Эти гены являются основными источниками зашиты пшеницы от листовой ржавчины, и их можно использовать в селекции на иммунитет. Кроме того, выделенные агрессивные патотипы KHPF, TRPG и FKLM позволяют использовать в иммунологических исследованиях для отбора сортов пшеницы на устойчивость к патогену в период проростков и взрослого растения.
Литература
- Кольбин Д.А., Волкова Г.В. Сорта зарубежной селекции, как источники неспецифической устойчивости к бурой ржавчине пшеницы // Материалы научно-практической конференции, посвященной 50-летию ВНИИБЗР. – Краснодар, 2010. – С.559-562.
- Койшибаев М. Болезни зерновых культур. – Алматы: Бастау, 2002. – 367 с.
- Рсалиев Ш.С., Койшибаев М.К., Моргунов А.И., Колмер Д. Анализ состава популяций стеблевой и листовой ржавчины пшеницы на территории Казахстана // Материалы международной научно-практической конференции. Алматы: Алейрон, 2005. – С. 267-272.
- Неттевич Э.Д. Рождение и жизнь сорта. – Москва: Московский рабочий, 1978. – 175 с.
- Макаров А.А., Коваленко Е.Д., Соломатин Д.А., Моторина Н.М. Методы полевой и лабораторной оценки неспецифической устойчивости растений к болезням // Материалы научного семинара «Типы устойчивости растений к болезням». – Санкт-Петербург, 2003. – С. 17-24.