Изучены сезонные изменения зараженности озерной лягушки Pelophylax ridibundus (Pallas) Девечинского лимана тремя видами трипаносом (Trypanosoma loricatum, T. neveulemairei и T. pipientis) в течение 2002 года. Приводятся показатели зараженности лягушек как отдельными видами жгутиконосцев, так и всеми тремя видами вместе. Установлено, что основным фактором, определяющим инвазированность амфибий кровепаразитами, является температура воды.
Одним из важных вопросов экологической паразитологии является изучение зависимости паразитофауны животных от сезонных явлений. Изучение влияния на паразитов сезонных изменений природных явлений имеет как теоретическое, так и практическое значение. Исследование сезонной периодичности инвазии животных паразитами, с одной стороны, дает материал для выяснения особенностей биологии и экологии паразитов, их взаимоотношений с хозяевами, с другой стороны, позволяет выявить как периоды наибольшей зараженности животных, а потому и наиболее опасные в эпизоотологическом отношении, так и моменты, когда наиболее целесообразно организовывать борьбу с патогенными видами.
Работ, посвященных изучению сезонных изменений зараженности рыб кровепаразитами, достаточно много (Ивасик, 1953; Залевская-Шаповал, 1954; Хайбулаев, 1969; Brumpt, 1905; Ahmed, 2002; Zintl, Poole, Voorheis et al., 2004; Hansen, 2005; Cottrell, 2007; Akmirza, Tepecik, 2007 и др.). Однако исследования по изучению сезонных изменений зараженности амфибий паразитами крови до нас нигде не проводились. В связи с этим целью настоящей работы явилось изучение сезонной динамики зараженности амфибий кровепаразитами.
При выборе объекта сезонных исследований зараженности амфибий кровепаразитами мы исходили из следующих соображений: выбранный нами вид амфибий не должен был совершать значительные миграции; быть достаточно многочисленным и легкодоступным; не представлять высокой коммерческой ценности; быть зараженным не одним, а хотя бы двумя или тремя видами кровепаразитов; иметь достаточно высокие показатели экстенсивности и интенсивности заражения кровепаразитами.
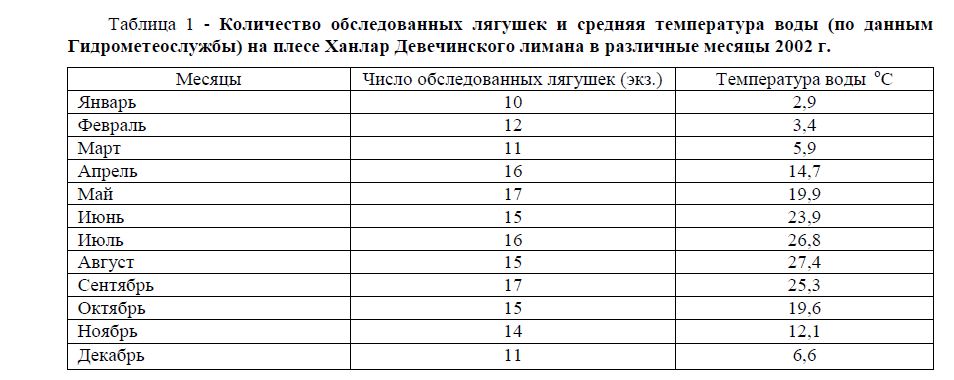
Всем указанным требованиям отвечала озерная лягушка Pelophylax ridibundus, обитающая в Девечинском лимане. Учитывая это, в 2002 году на плесе Ханлар Девечинского лимана мы проводили сезонные исследования зараженности озерной лягушки кровепаразитами. Исследования охватили период с января по декабрь (таблица 1). На наличие кровепаразитов было обследовано 169 особей озерной лягушки. Чтобы сезонные исследования охватили животных примерно одного возраста, мы вскрывали лягушек размером от 30 мм до 60 мм, которые состояли из сеголеток и годовиков.
Мазки, приготовленные из периферической крови лягушек, фиксировали 96°-ным этанолом и окрашивали по Романовскому-Гимза (рН 7,2).
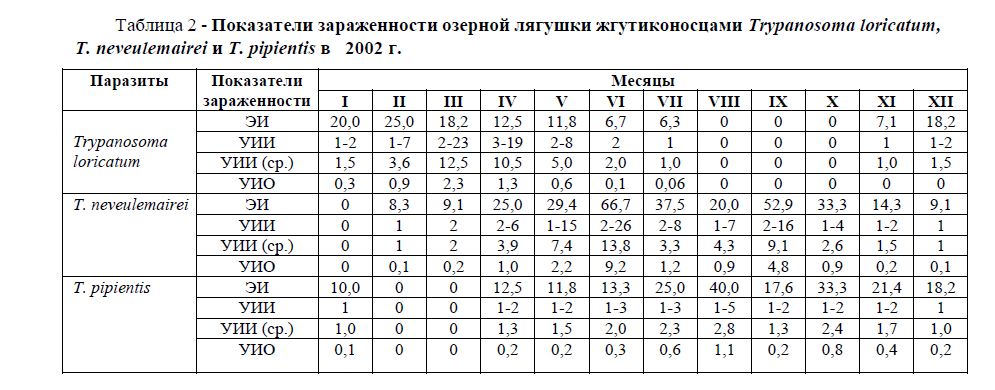
В таблице 2 приводятся показатели экстенсивности инвазии – ЭИ (%), удельной интенсивности инвазии – УИИ (количество паразитов, приходящихся на мазок крови), средней удельной интенсивности инвазии – УИИ (ср.) и удельный индекс обилия – УИО (сумма паразитов, обнаруженных в одном мазке каждого зараженного хозяина, разделенная на количество исследованных хозяев) каждого паразита за каждый месяц года. Более наглядно эти данные приведены в рисунках 1 и 2.
Таблица 1 - Количество обследованных лягушек и средняя температура воды (по данным Гидрометеослужбы) на плесе Ханлар Девечинского лимана в различные месяцы 2002 г.
По данным наших исследований, в самом начале января ЭИ жгутиконосцем Trypanosoma loricatum составила 20,0%; УИИ был невысокий – 1-2 экз., УИИ (ср.) – 1,5 экз. В феврале ЭИ изменяется незначительно, она поднимается до 25,0%, в то же время наблюдается изменение удельной интенсивности инвазии (1-7 экз.), УИИ (ср.) повышается более чем в два раза (3,6 экз.). В марте УИИ продолжает расти - 2-23 экз.; УИИ (ср.) - 12,5 экз., несмотря на то что экстенсивность инвазии даже несколько понижается (до 18,2%). Это свидетельствует о том, что, возможно, происходит размножение T. loricatum в организме лягушек. В апреле начинается понижение зараженности лягушек жгутиконосцем T. loricatum: ЭИ - 12,5%; УИИ - 3-19 экз.; УИИ (ср.) - 10,5 экз.), которая продолжается в мае: ЭИ - 11,8%; УИИ - 2-8 экз.; УИИ (ср.) - 5,0 экз., июне: ЭИ - 6,7%; УИИ - 2 экз.; УИИ (ср.) - 2,0 экз. и июле: ЭИ - 6,3%; УИИ - 2 экз.; УИИ (ср.) - 1,0 экз. В августе, сентябре и октябре жгутиконосца T. loricatum у озерной лягушки мы ни разу не отметили. В ноябре этот паразит отмечен в крови одной лягушки: ЭИ - 7,1%; УИИ - 1 экз.; УИИ (ср.) - 1,0 экз., а в декабре – в крови двух лягушек: ЭИ - 18,2%; УИИ - 1-2 экз.; УИИ (ср.) - 1,5 экз.
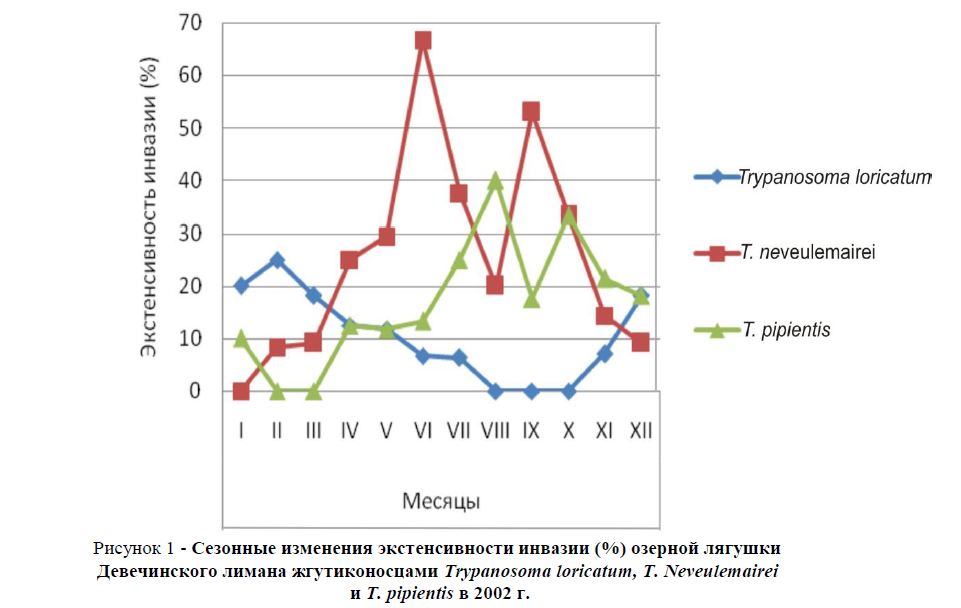
Рисунок 1 - Сезонные изменения экстенсивности инвазии (%) озерной лягушки Девечинского лимана жгутиконосцами Trypanosoma loricatum, T. Neveulemairei и T. pipientis в 2002 г.
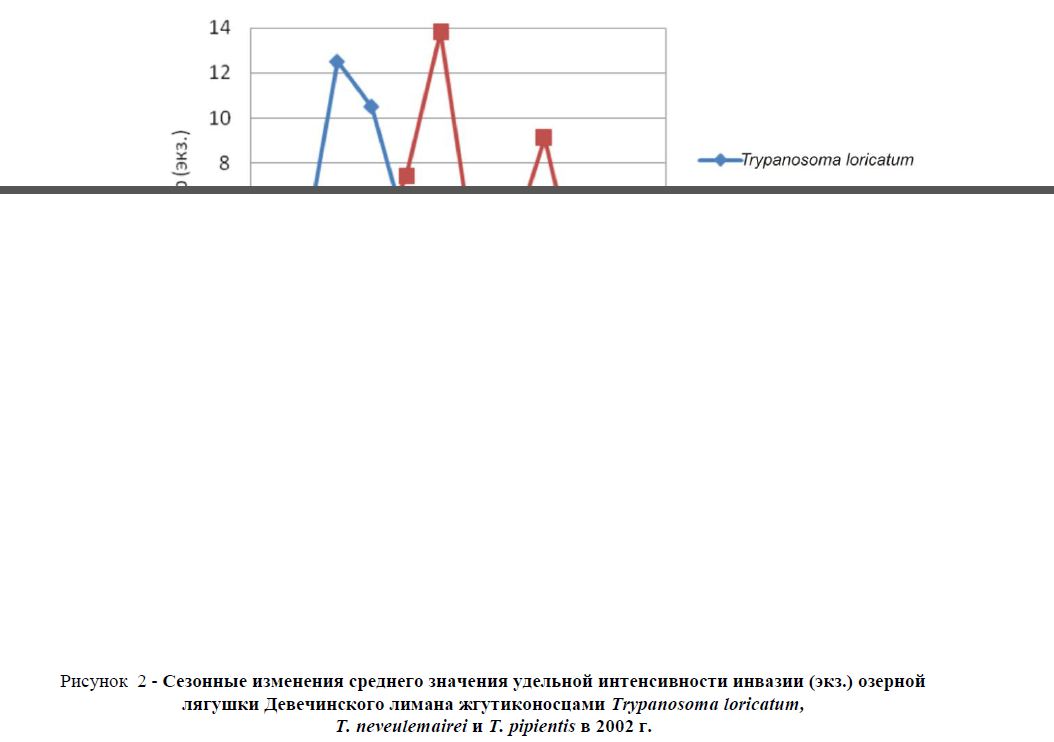
Рисунок 2 - Сезонные изменения среднего значения удельной интенсивности инвазии (экз.) озерной лягушки Девечинского лимана жгутиконосцами Trypanosoma loricatum,neveulemairei и T. pipientis в 2002 г.
О характере изменений зараженности озерной лягушки жгутиконосцем T. loricatum можно сказать следующее: зимой и в начале лета происходит заражение лягушек этим паразитом, весной и в начале лета, по-видимому, трипаносомы размножаются в организме лягушек, а к концу лета зараженность сильно падает. Единичные особи паразита начинают появляться в крови лягушек осенью.
В январе жгутиконосец T. neveulemairei в крови лягушек не отмечен. В феврале и в марте он найден по одному разу: ЭИ – соответственно, 8,3% и 9,1%; УИИ - 1 экз. и 2 экз.; УИИ (ср.) - 1,0 экз. и 2,0 экз. В апреле с повышением температуры воды начинается заметное увеличение показателей зараженности, ЭИ составляет 25,0%, УИИ - 3-6 экз., УИИ (ср.) - 3,9 экз. В мае это повышение зараженности продолжается: ЭИ достигает 29,4%, УИИ - 1-15 экз., УИИ (ср.) - 7,4 экз. В июне зараженность достигает максимума, ЭИ доходит до 66,7%, УИИ составляет 2-26 экз., УИИ (ср.) - 13,8 экз. В июле происходит падение зараженности, ЭИ понижается до 37,5%, УИИ - до 2-8 экз., а УИИ (ср.) - до 3,3 экз. В августе понижение зараженности продолжается: ЭИ составляет 20,0%, УИИ - 1-7 экз., УИИ (ср.) - 4,3 экз. В сентябре, когда температура воды еще достаточно высокая для размножения кровепаразитов, ЭИ равнялась 52,9%, УИИ - 2-16 экз., УИИ (ср.) - 9,1 экз. В дальнейшем, в связи с падением температуры воды, зараженность лягушек этим паразитом понижается: в октябре ЭИ уменьшается до 33,3%, УИИ - до 1-4 экз., а УИИ (ср.) - до 2,6 экз.; в ноябре ЭИ падает еще до 14,3%, УИИ - до 1-2 экз., а УИИ (ср.) - до 1,5 экз.; в декабре ЭИ падает до 9,1%, УИИ - до 1 экз., а УИИ (ср.) - до 1 экз.
По приведенным выше данным, в 2002 году экстенсивность инвазии и интенсивность инвазии жгутиконосцем T. neveulemairei, который заражает озерную лягушку Девечинского лимана сильнее других кровепаразитов в течение года, совершают два пика повышения и два пика понижения, образуя двухвершинные кривые. Первая вершина каждой из этих кривых приходятся на июнь, а вторая вершина - на сентябрь месяцы.
Третий из кровепаразитов озерной лягушки T. pipientis, сезонную динамику зараженности которого мы исследовали, в январе найден в крови одной из десяти (10,0%) исследованных лягушек. УИИ при этом составила 1 экз., УИИ (ср.) – 1,0 экз.
В феврале и марте среди обследованных лягушек не было зарегистрировано особей, зараженных жгутиконосцем T. pipientis. В каждый из месяцев (апрель, май и июнь) было зарегистрировано по две особи лягушек, зараженных этим паразитом: ЭИ была, соответственно, 12,5%, 11,8% и 13,3%. УИИ составляла при этом 1-2 экз., 1-2 экз. и 1-3 экз., а УИИ (ср.) - 1,3 экз., 1,5 экз. и 2,0 экз. В июле начинается повышение зараженности озерной лягушки жгутиконосцем T. pipientis. Это повышение продолжается два месяца, в июле ЭИ составляет 25,0%, УИИ - 1-3 экз., а УИИ (ср.) - 2,3 экз., в августе ЭИ достигает 40,0%, УИИ - 1-5 экз., а УИИ (ср.) - 2,8 экз. В сентябре ЭИ падает до 17,6%, УИИ - до 1-2 экз., а УИИ (ср.)
- до 1,3 экз. В октябре происходит небольшое повышение зараженности, ЭИ доходит до 33,3%, УИИ остается 1-2 экз., а УИИ (ср.) повышается до 2,4 экз. В ноябре и декабре происходит понижении заражения: ЭИ составляет, соответственно, 21,4% и 18,2%, УИИ - 1-2 экз. и 1 экз., УИИ (ср.) - 1,7 экз. и 1 экз.
Как экстенсивность, так и интенсивность инвазии озерной лягушки жгутиконосцем T. pipientis медленно повышается с марта по август, в сентябре немного понижается, в октябре чуть повышается и далее до конца года идет на понижение.
Сравнение сезонных изменений зараженности озерной лягушки Девечинского лимана тремя видами трипаносом, которое представлениа на рисунках 1 и 2, показывает следующее:
В январе УИИ (ср.) озерной лягушки видами Trypanosoma loricatum (1,5 экз.) и T. pipientis (1,0 экз.) была низкой, а ЭИ составляла, соответственно, 20,0% и 10,0%. T. neveulemairei в январе не была отмечена.
В феврале ЭИ и ИИ видом T. loricatum несколько повышается, появляется вид T. neveulemairei, а вид
- T. pipientis исчезает.
В марте отмечены только T. loricatum и T. neveulemairei. ЭИ первым повышается немного, а ИИ повышается очень существенно и достигает пика.
В апреле продолжается повышение ЭИ и ИИ видом T. neveulemairei. У вида T. loricatum эти показатели немного понижаются. Появляется вид T. pipientis, но зараженность им невысокая.
В мае ЭИ и ИИ видом T. neveulemairei. заметно повышаются, а эти же показатели инвазии видом
- T. loricatum - понижаются. Зараженность видом T. pipientis почти не изменяется, оставаясь невысокой.
В июне ЭИ и ИИ видом T. neveulemairei достигает максимума, зараженность видом T. loricatum
понижается, а видом T. pipientis почти не изменяется, оставаясь как и в мае невысокой.
В июле ЭИ и ИИ видом T. neveulemairei резко падают, в это же время происходит резкое повышение тех же показателей по T. pipientis. Экстенсивность инвазии видом T. loricatum не изменяется, а интенсивность понижается.
В августе ЭИ видом T. neveulemairei падает еще ниже, а ИИ изменяется незначительно. В этот же месяц ЭИ и ИИ видом T. pipientis повышается, достигнув своего максимума. T. loricatum исчезает и не появляется до ноября.
В сентябре ЭИ и ИИ видом T. neveulemairei снова резко повышается, образуя второй пик.
Одновременно падает ЭИ и ИИ видом T. pipientis.
В октябре ЭИ и ИИ видом T. neveulemairei падают, в то же время резко повышаются оба эти показатели по виду T. pipientis. Создается впечатление, что между этими видами существует антагонизм, не позволяющий одновременное повышение заражения лягушки обоими из них. В ноябре зараженность видами T. neveulemairei и T. pipientis падает, причиной может быть понижение температуры воды. В это же время в крови лягушек появляется вид T. loricatum. Однако зараженность им бывает невысокой. В декабре зараженность видами T. neveulemairei и T. pipientis продолжает понижаться, а зараженность видом T. loricatum несколько повышается.
Из совместного анализа изменений зараженности озерной лягушки Девечинского лимана всеми тремя видами жгутиконосцев видно, что зараженность видом T. loricatum в течение всего года остается невысокой.
Таким образом, исследования сезонных изменений зараженности озерной лягушки, проведенные нами в 2002 году, показали, что основным фактором, определяющим прямо и косвенно инвазированность амфибий кровепаразитами, является температура воды. Прямое воздействие температуры воды заключается в стимулировании размножения жгутиконосцев, которое активизируется при температуре воды между 15-16ОС. Резкое повышение интенсивности инвазии кровепаразитами вызывает повышение резистентности организма лягушек в ответ на гиперинвазию, в результате этого зараженность снижается. После этого резистентность организма лягушек несколько падает и, при наличии благоприятных температурных условий, происходит повторное повышение интенсивности и экстенсивности инвазии. Опосредованное же воздействие температурного фактора выражается в том, что активное нападение пиявок, являющихся переносчиками кровепаразитов, на лягушек, а также размножение жгутиконосцев в пиявках приходится, в основном, на холодное время года.
Таблица 2 - Показатели зараженности озерной лягушки жгутиконосцами Trypanosoma loricatum, neveulemairei и T. pipientis в 2002 г.
Литература
- Ивасик В.М. Паразиты карпа в рыбхозах западных областей УССР и болезни, ими вызываемые // Труды НИИПОРХ УССР. - Киев, 1953. - Т. 9. - С. 85-122.
- Залевская-Шаповал Н.М. Кровепаразитические простейшие рыб бассейна Днепра: автореф. дис. … канд. биол. наук. – Киев, 1954. - 20 с.
- Хайбулаев К.Х. Кровепаразитические простейшие рыб Каспийского моря: дис. … канд. биол. наук. – Махачкала, 1969, - 232 с.
- Brumpt E. Trypanosomes et trypanoplasmes. Rev. Sci. - Paris, 1905. - N. 4. - 165
- Ahmed Sh.M. A Study on Trypanosomiasis in some Freshwater Fishes at Assiut Envir. Encyclopedia Ass. Univ., 2002. - Vol. 45. - No. 2. - Р. 5-17.
- Zintl , Poole W.R., Voorheis H.P., Holland C.V. Naturally occurring Trypanosoma granulosum infections in the European eel, Anguilla anguilla L. from County Mayo, western Ireland. // Journal of Fish Disease. - 2004. - Vol. 27. - Issue 5. - Р. 333-341.
- Hansen Infection of haemoflagellates (Protozoa, Kinetoplastida) in freshwater fish from the lake Vannsjo, Norway. Cand. Scient. / Thesis in Parasitology, University of Oslo, Zoological Museum, Sars gate 1, N-0562.
- - Oslo, 2005. - 44 p.
- Cottrell J. A trypanosome from the plaice, Pleuronectes platessa L. // Journal of Fish Biolog. - 2007. - Vol. 11 21 Issue 1. - Р. 35 – 47.
- Akmirza , Tepecik R. Seasonal variation in some haematological parameters in naturally infected and uninfected roach (Rutilus rutilus) with Cryptobia tincae. // Journal of Applied Biological Sciences. - 2007. - Vol. 1. - No. 3. - Р. 61-65.