Изучение редких и исчезающих растений регионов Казахстана имеет важное практическое значение для сохранения биологического разнообразия и мониторинга состояния растительности. Одним из показателей состояния популяций является изучение структуры фаз развития растений. Цель исследования — проведение исследования этапов онтогенеза Pulsatilla uralensis (Zam.) Tzvel. на территории Северного Казахстана. В статье приведены итоги изучения возрастных особенностей онтогенеза и структуры ценопопуляций Pulsatilla uralensis на территории Костанайской области. Ценопопуляции P. uralensis приурочены к лесным опушкам или вторичным степным сообществам на месте старых гарей на почвах легкого механического состава. В онтогенезе выделено три периода и 8 возрастных состояний: проростки, ювенильное, имматурное, виргинильное, молодое, средневозрастное, старовозрастное генеративное, субсенильное состояния. Особенностью возрастных состояний P. uralensis по сравнению с сибирскими и якутскими популяциями является более ранее разделение корневищ на отдельные парцеллы с образованием рыхлого каудекса, наблюдаемое в средневозрастном генеративном состоянии. В разреженном сосновом лесу формируется нормальная популяция с преобладанием генеративных особей, относящаяся к зрелым популяциям с низким индексом восстановления. В популяциях, сформированных в условиях вторичной псаммофитной степи на месте сгоревшего соснового леса, произрастает левосторонняя инвазионная популяция с преобладанием имматурных и виргиниль- ных особей, обладающая высоким индексом восстановления.
Введение
Изучение популяций редких и исчезающих растений в Казахстане необходимо для понимания современного их состояния, организации мониторинга и мероприятий по охране данных видов. Среди оценочных показателей важную роль играют онтогенетические исследования, так как изучение соотношения фаз онтогенеза является важным показателем не только современного состояния популяции растения, но и динамики развития в будущем.
В «Красной книге Казахстана» [1] приводится Pulsatilla flavescens (Zucc.) Juz. Эту комбинацию предложил С.В. Юзепчук (1937) во «Флоре СССР», но оказалось, что видовой эпитет «flavescens» в роде Pulsatilla уже был использован для другого вида, в этом случае «Кодексом ботанической номенклатуры» [2] предусматривается создание новой номенклатурной комбинации. Н.Н. Цвелев [3] предложил использовать название Pulsatilla uralensis (Zam.) Tzvel., для которого характерны околоцветники желтой гаммы окраски, узкие клиновидные к основанию листочки, черешок у верхней доли или очень короткий, или его нет.
Распространён P. uralensis в европейской части России, а именно Волжско-Камском районе, Южном Урале [3–5]. В Казахстане он встречается в Тобол-Ишимском, Семипалатинском боровом, Кокчетавском флористических районах, на севере Западного мелкосопочника и Тургайского флористического района [6]. Ценоареалы P. uralensis, в основном, приурочены к степным, опушечным фитоценозам на почвах легкого механического состава [3–5]. Особенностью Костанайской области является то, что P. uralensis с небольшим обилием встречается в сосновых лесах на песках и супесях [7]. Несмотря на довольно широкий ареал P. uralensis, встречается довольно редко, хозяйственное освоение территорий и, прежде всего, распашка земель привели к тому, что в Казахстане и некоторых субъектах Российской Федерации этот вид включен в «Красные книги» [1, 8–10]. Несмотря на редкость вида, сокращение численности популяций исследований биологических особенностей P. uralensis на территории Казахстана не проводилось.
Целью наших исследований являлось изучение онтогенеза и структуры популяций P. uralensis на территории Костанайской области.
Объекты и методика исследований
Объектом исследования были ценопопуляции P. uralensis, расположенные в Костанайской области. Для изучения ценопопуляций (ЦП) в мае 2018 и 2019 гг. подбирались участки с высокой плотностью цветущих особей. О.Е. Сушенцов [11], изучавший структуру популяций P. uralensis и P. patens на Урале, на большом статистическом материале показал, что между двумя этими видами образуется «серая зона» в виде переходных популяций, обладающих промежуточными значениями признаков. Поэтому выбирались только те ценопопуляции, в которых желтоцветковых особей было 100 %. Проводилось флористическое описание ЦП с указанием количества видов, общего и парциального проективного покрытия P. uralensis.
Границы популяции определяли общепринятыми методами [12], площадь изучалась с помощью GPS. Флористические описания проводились стандартным методом на площади 100 м2, оценивались видовой состав, общее и частное проективное покрытие каждого вида. Внутри популяции закладывалось 10 модельных площадок по 1 м2, на которых подсчитывалось количество особей всех возрастных состояний. В качестве счетной единицы использовали особь P. uralensis любого возрастного состояния. Возрастные состояния выделены, согласно методическим указаниям Т.А. Работнова [13], А.А. Уранова [14], О.В. Смирновой и других [12]. Поскольку проростки и ювенильные растения в полевых условиях найти не удалось, они описывались по экземплярам, полученным при посеве свежесобранными семенами. Тип ценопопуляции определяли по Т.А. Работнову [13], классификацию популяций по «дельта-омега» по Л.А. Животовскому [15].
Индекс возрастности ценопопуляции (Δ) вычисляется по формуле
Δ = ΣKimi ⁄ ΣKi,
где ΣKi — сумма растений всех возрастных состояний; mi — возрастность особей [15].
Индекс эффективности (ω) определяли следующим образом:
ω = Σ piei,
где pi = ni /n — доля растений i-того состояния в данной популяции; ni — абсолютное число растений i-того состояния, n = Σni — общее число растений; ei — энергетическая эффективность.
Индекс восстановления (I) определялся по формуле
I = Σj→v ⁄ Σ g1→g3, где Σj→v — сумма растений всех возрастных состояний прегенеративного периода; Σg1→g3 — сумма растений всех возрастных состояний генеративного периода [16].
Результаты и обсуждение
Латентный период. Плод P. uralensis — многоорешек. Каждый орешек удлиненно-конический, заостренный, 3–4 мм длиной, с волосистой остью 15–20 мм дл. Созревание орешков происходит в конце мая. В середине июня они осыпаются. Орешки, снабженные остью, могут планировать на некоторое расстояние от материнского растения. На влажной почве они втыкаются в почву и в результате раскручивания ости ввинчиваются в почву на глубину до 2 см. Семенная продуктивность 140–170 орешков на плод. Реальная семенная продуктивность составляет в природных условиях 500–600 орешков, что согласуется с результатами, полученными у близкородственных видов [17, 18] (рис. 1).
Виргинильный период. Проростки (p). Семена не имеют периода покоя, они могут прорастать осенью, но чаще всего прорастают в конце апреля следующего года. Грунтовая всхожесть высокая, около 50 %, лабораторная — 85 %. Прорастание надземное, семядоли продолговатые короткочерешковые, из пазухи которых появляется двулопастной первый настоящий лист. Ниже семядолей располагается гипокотиль, переходящий в первичный стержневой корешок. В состоянии проростков формируется розеточный побег из 3–5 листочков разной сложности (рис. 2, p).
В ювенильном состоянии (j) формируется розеточный побег из 4–5(6) листьев, в основании которых закладываются почки возобновления. Листья трехраздельные, рассеченные на крупные доли, которые, в свою очередь, надрезаны на зубцы. Корневая система представлена стержневым корнем и многочисленными корнями второго порядка (рис. 2, j).
Е.А. Сивцева [19] обозначает жизненную форму P. uralensis в этих возрастных состояниях как растения со стержневой корневой системой, вегетативным моноподиально нарастающим розеточным побегом.
У имматурных особей (im) происходит формирование удлиненного корневища, с сохранением остатков прошлогодних листьев (по числу которых обычно определяется календарный возраст растений [20]. Сохраняется стержневой корень, который ветвится на глубине 5–7 см на корни второго порядка. Из почек возобновления, находящихся в пазухе нижних настоящих листьев, образуется 1–3 розеточных побегов. Листья трехраздельные, доли первого порядка трехлопастные (рис. 2, im).
Рисунок 3. Возрастные состояния P. uralensis: v — виргинильное состояние
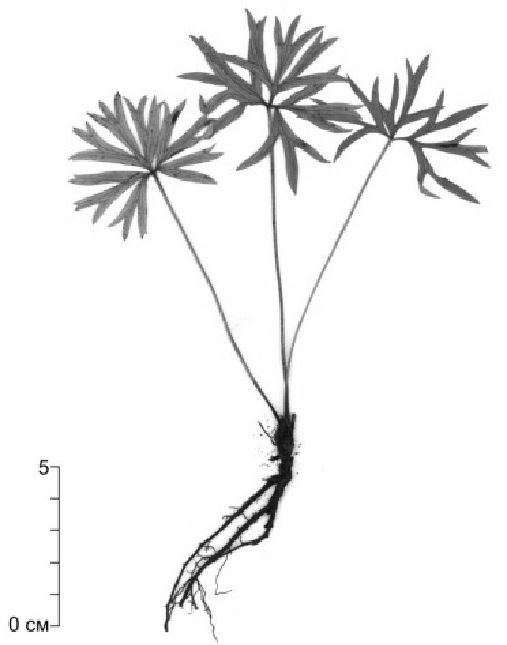


В виргинильном состоянии (v) происходит нарастание количества почек возобновления и розеточных модулей Н.А. Цыбанова [20] называет их узлами кущения), формирование вертикального корневища с многочисленными почками возобновления. У молодых виргинильных особей насчитывается 4–6 шт. листьев, на поздних стадиях их количество возрастает до 9–12 шт. (рис. 3, v).
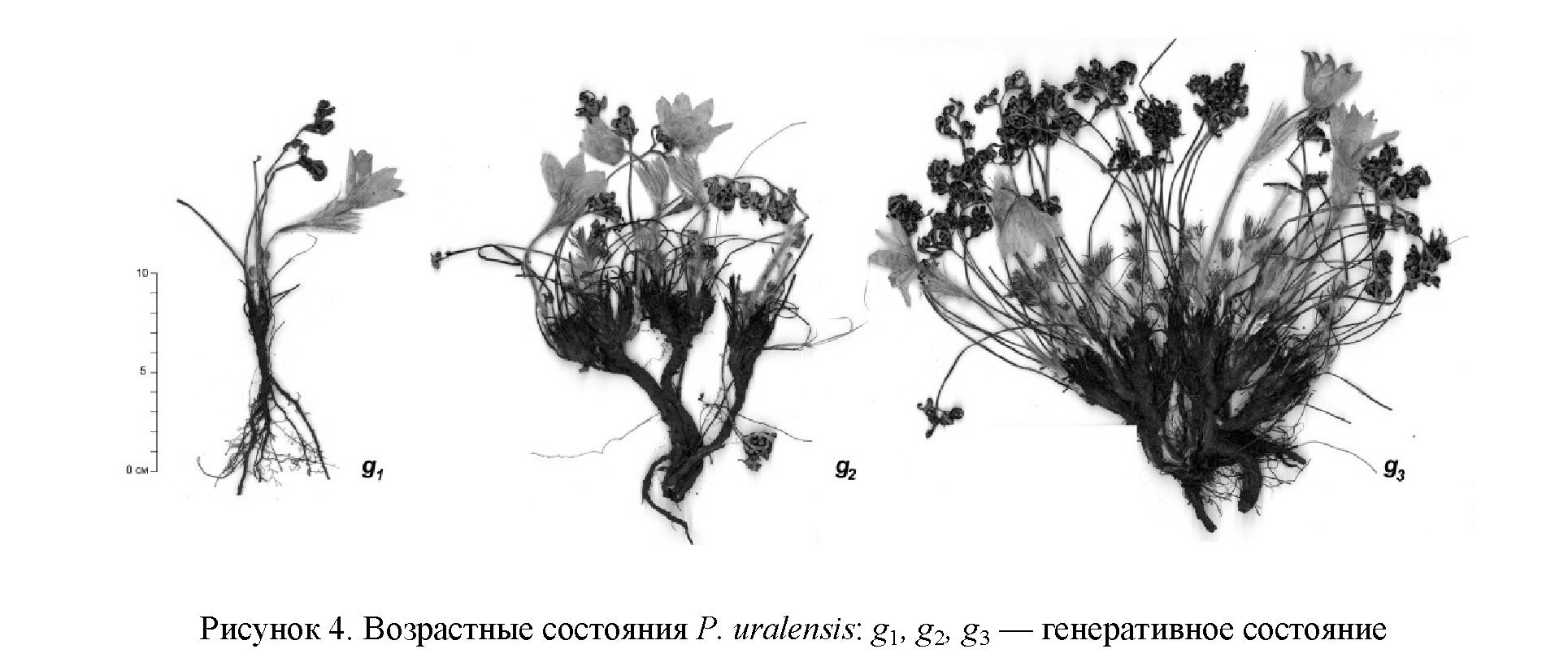
Генеративный период. Первые цветки, характеризирующие переход растений в молодое генеративное состояние (g1), появляются из верхушечной почки, из боковых пазушных почек, к концу цветения генеративного побега появляются 1–2 вегетативных розеточных побега. В молодом генеративном состоянии образуются 1–2 цветущих побега, высотой 15–17 см, при плодах увеличивается до 23– 28 см. В этом возрасте отмечено разветвление корневища (рис. 4, g1).
В среднем генеративном состоянии (g2) корневище представлено многоглавым каудексом, продолжается его интенсивное ветвление на отдельные парцеллы. В отличие от P. uralensis и Pulsatilla multifida (G. Printz.) Juz., в Якутии [19] ветвление корневищ происходит на глубине до 10 см, образуя несколько парциальных модулей. Каждый из них несет 1–3 генеративных побегов и 4–6 розеточных побегов, образующих рыхлую дернину (рис. 4, g2). В этом возрасте растение остается вегетативно малоподвижным, и мы не наблюдали корневых отпрысков, обнаруженных у P. patens [20].
У старовозрастных генеративных особей (g3) происходит полная или частичная мацерация корневища с образованием 3–6 парциальных кустов I-го порядка, которые образуют парциальные кусты II-го порядка. Образование вегетативных розеточных побегов продолжается, но снижается количество генеративных побегов, которые образуются не в каждом парциальном кусте (рис. 4, g3).
Субсенильные особи (s) представлены системой отмирающих корневищ, с одиночными вегетативными розетками. Листовые пластинки сохраняют видовую специфичность, но они более мелкие, количество зубцов на листьях уменьшается (рис. 5, s).
Необходимо отметить, что имеются некоторые отличия возрастных состояний популяций
P. uralensis в Северном Казахстане от якутских популяций [19]. Они касаются более раннего (g2) разделения корневищ на отдельные парцеллы с образованием рыхлого каудекса.
По классификации жизненных форм И.Г. Серебрякова [21], P. uralensis является многолетним травянистым коротко-стержневым многоглавым поликарпиком. В отличие от других видов, Pulsatilla Mill. P. uralensis характеризуется парциальной системой побегов, с разделением корневищ на значительной глубине почв.
Изучена возрастная структура двух ЦП P. uralensis, расположенных на территории Костанайской области:
ЦП-1. Костанайская обл., окр. с. Озерное, Новонеженское л-во, кв. 21, N52.43971°, E64.09279°, h = 213 м н. у. м.; разреженный сосновый лес на песках. Площадь популяции — 2000 м2, ОПП — 40 %, ОПП P. uralensis — 20 %, сообщество насчитывает 21 вид. В сложении растительности наибольшее участие имеют: Agropyron cristatum, Betula pendula, Centaurea ruthenica, Cerasus fruticosa, Eremogone longifolia, Festuca beckeri, Jurinea cyanoides, Otites wolgensis, Pinus sylvestris, Populus tremula, Ranunculus polyanthemos, Stellaria graminea.
ЦП-2. Костанайская обл., окр. с. Щербаково, N53.20453°, E64.21550°, h = 193 м н. у. м.; вторичная псаммофитная степь на месте сгоревшего соснового леса. Площадь популяции — 3000 м2, ОПП — 100 %, ОПП P. uralensis — 10 %, сообщество насчитывает 27 видов. В сложении растительности наибольшее участие имеют: Achillea millefolium, A.setacea, Artemisia campestris, A. scoparia, Bromopsis inermis, Calamagrostis epigeios, Conioselinum tataricum, Helichrysum arenarium, Koeleria glauca, Melilotus officinalis.
Ценопопуляции P. uralensis приурочены к лесным опушкам или вторичным степным сообществам на месте старых гарей на почвах легкого механического состава. Популяции не очень большие по площади, 2–3 тыс. шт./100 м2, их плотность 5–6 шт./м2 (см. табл.), что вполне соответствует норме для корнестержневых многолетников [22].
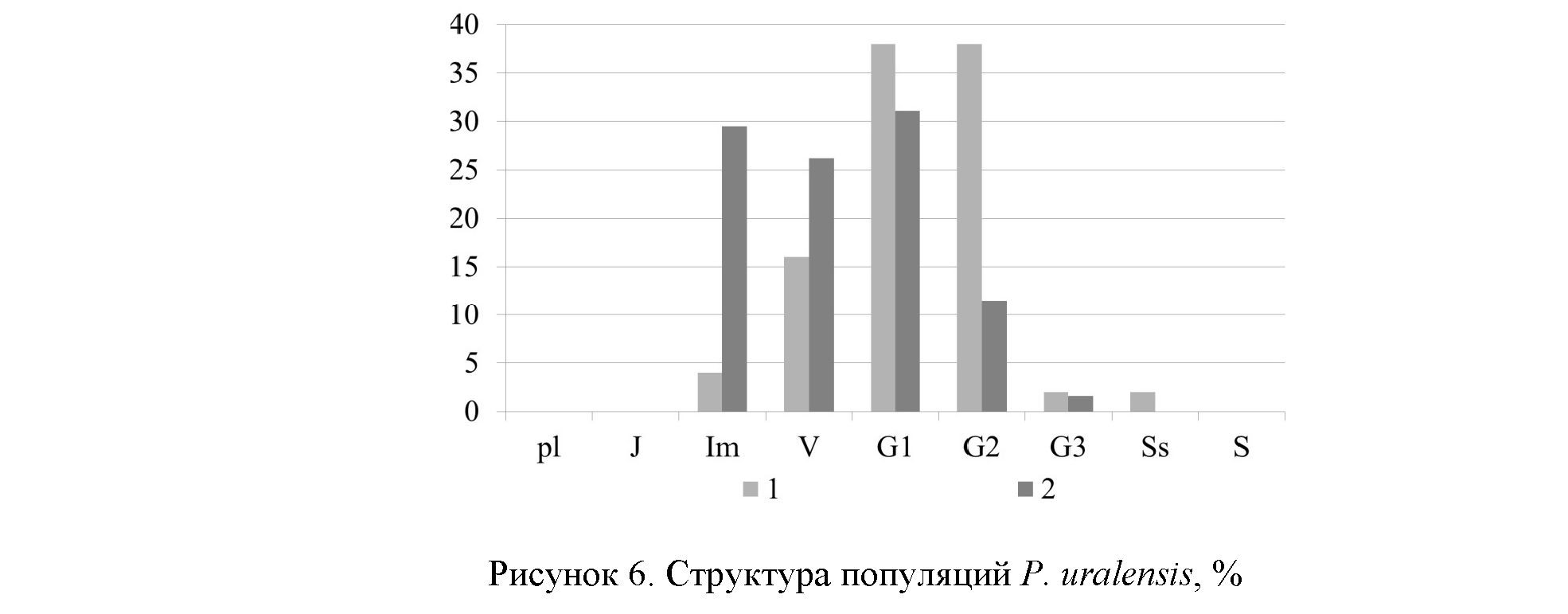
В возрастных спектрах ЦП-1 наибольшая доля принадлежит генеративным особям, поэтому спектры правосторонние, одновершинные с максимумами на молодых и средневозрастных генеративных растениях (рис. 6), что вполне согласуется с результатами, полученными для других видов Pulsatilla [23].
Таблица Характеристика ценопопуляций P. uralensis
|
№ ЦП |
ОПП, % |
ОПП P. uralensis, % |
Площадь ЦП, м2 |
Плотность особей, шт./1 м2 |
Количество особей в ЦП, шт. |
Δ |
Ω |
I |
|
ЦП-1 |
40 |
20 |
2000 |
6,10 |
12,2×103 |
0,54 |
0,83 |
0,16 |
|
ЦП-2 |
100 |
10 |
3000 |
5,00 |
15×103 |
0,35 |
0,74 |
0,53 |
В ЦП-2 доля имматурных и виргинильных особей составляет более 50 %, и возрастной спектр становится левосторонним, что связано с тем, что популяция сформировалась на вторичном экотопе и в отсутствии конкурентности со стороны других растений. В ценопопуляциях длительно живущих стержнекорневых многолетников могут доминировать средневозрастные генеративные особи в связи с наибольшей продолжительностью этого периода и наименьшей элиминацией в это время [24]. Всходы и ювенильные особи в популяциях не обнаружены (рис. 6).
Доля постгенеративных растений незначительна, что неоднократно отмечалось у стержнекорневых травянистых поликарпиков [23, 24].
Индекс возрастности оценивает онтогенетический уровень ЦП в конкретный момент времени. Он может иметь значения от 0 до 1,0. Чем выше его показатель, тем старше исследуемая ценопопу- ляция [14, 15]. Возрастность в ЦП-1 соответствует зрелым популяциям, а в ЦП-2 — молодым. Индекс эффективности (энергетическая нагрузка на среду, вызываемая «средним» растением [15]), в ЦП-1 составляет 0,83, в ЦП-2 — 0 ,74. По классификации популяций «дельта — омега» обе популяции относятся к зрелым.
145
Индекс восстановления определяется содержанием в популяциях доли особей прегенеративного периода и изменяется от 0 до 1,0. В ЦП-1 он очень низок и составляет 0,16, это свидетельствует о старении популяции. В ЦП-2 он достаточно высок — 0,53, что говорит об инвазионном характере популяции и ее неустойчивости.
Заключение
Pulsatilla uralensis (Zam.) Tzvel. (=Pulsatilla flavescens (Zucc.) Juz.) относится к многолетним травянистым коротко-стержневым многоглавым поликарпикам. Он распространён в европейской части России (Волжско-Камский район, Южный Урал) а также в северо-западных областях Казахстана. Растения встречаются в степной зоне, местообитания приурочены к степным травянистым сообществам, степным борам и опушкам леса. Особенностью возрастных состояний P. uralensis в Северном Казахстане является разделение корневищ на глубине 10 см с образованием нескольких парциальных модулей и формированием рыхлого каудекса с полициклическими побегами.
На структуру ценопопуляций большое влияние оказывают условия формирования. В разреженном сосновом лесу оказывается нормальная популяция с преобладанием генеративных особей, относящаяся к зрелым популяциям с низким индексом восстановления.
В популяциях, сформированных в условиях вторичной псаммофитной степи на месте сгоревшего соснового леса, наблюдается левосторонняя инвазионная популяция с преобладанием имматурных и виргинильных особей, обладающая высоким индексом восстановления.
Результаты данных исследований могут использоваться для мониторинга степной растительности на особо охраняемых природных территориях Костанайской области.
Работа выполнялась в рамках Проекта грантового финансирования Министерства образования и науки Республики Казахстан на 2018–2020 гг. № AP05132458 «Молекулярно-генетический анализ генофондов популяций редких видов растений Северного Казахстана» ( номер гос. регистрации 0118РК00404 и гос. задания № 0352–2016–0002 «Оценка состояния и охрана флористического разнообразия под влиянием антропогенных и техногенных факторов in situ и ex situ»; USU 508667 «Гербарий КузБС» (KUZ)).
Список литературы
- Красная книга Казахстана. — Т. 2. Растения / гл. ред. И.О. Байтулин; отв. ред. Г.Т. Ситпаева. — 2-е изд. перераб. и доп. — Астана: «Арт-Print XXI» Ltd, 2014 — 452 с.
- Международный кодекс ботанической номенклатуры (Венский кодекс). — М.; СПб., 2009. — 282 с.
- Цвелев Н.Н. Триба 7. Anemoneae DC / Н.Н. Цвелев // Флора Восточной Европы. — М.; СПб.: Мир и семья, 2001. — Т. 10. — С. 77–95.
- Куликов П.В. Определитель сосудистых растений Челябинской области / П.В. Куликов. — Екатеринбург: Ботсад УрО РАН, 2010. — 968 с.
- Науменко Н.И. Флора и растительность Южного Зауралья / Н.И. Науменко. — Курган: Изд-во Курган. гос. ун-та, 2008. — 511 с.
- Гамаюнова А.П. Род Прострел — Pulsatilla Mill. / А. П. Гамаюнова // Флора Казахстана. — Т. 4. — Алма- Ата, 1961. — С. 66–70.
- Пугачев П.Г. Сосновые леса Тургайской впадины / П. Г. Пугачев. — Кустанай, 1994. — 408 с.
Серия «Биология. Медицина. География». № 3(99)/2020
- Красная книга Ханты-Мансийского автономного округа — Югры. — Екатеринбург: Баско, 2013. — 460 с.
- Красная книга Свердловской области: Животные, растения, грибы. — Екатеринбург: Мир, 2018. — 450 с.
- Красная книга Республики Башкортостан: [В 2 т.]. — Т. 1: Растения и грибы. — Уфа: Медиа-Принт, 2011. — 384 с.
- Сушенцов О.Е. Структура популяций видов Pulsatilla (Ranunculaceae) на Урале / О.Е. Сушенцов // Бот. журн. — 2007. — Т. 92, № 4. — С. 493–505.
- Смирнова О.В. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений разных биоморф / О.В. Смирнова, Л.Б. Заугольнова, Н.А. Торопова, Л.Д. Фаликов // Ценопопуляции растений. (Основные понятия и структура). — М.: Наука, 1976. — С. 14–44.
- Работнов Т.А. Жизненный цикл многолетних травянистых растений в лесных ценозах / Т.А. Работнов // Тр. БИНа АН СССР. Сер. 3. — 1950. — Вып. 6. — С. 7–204.
- Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических процессов / А.А. Ура- нов // Биологические науки. — 1975. — № 2. — С. 7–34.
- Животовский Л.А. Онтогенетические спектры, эффективная плотность и классификация популяций растений / Л.А. Животовский // Экология. — 2001. — № 1. — С. 3–7.
- Жукова Л.А. Динамика популяций луговых растений в естественных фитоценозах / Л.А. Жукова // Динамика цено- популяций травянистых растений: сб. науч. тр. — Киев: Наук. думка, 1987. — С. 9–19.
- Косюкова О.В. Семенное размножение Pulsatilla patens (L.) Mill. и Pulsatilla pratensis (L.) Mill. / О.В. Косюкова, Ю.А. Демочко, Н.А. Петрова, И.В. Шилова // Бюл. Бот. сада Саратов. ун-та. — 2014. — Т. 12, Вып. 1. — С. 110–119.
- Панин А.В. Семенная продуктивность Pulsatilla patens (L.) Mill. и Pulsatilla pratensis (L.) Mill. в условиях г. Саратова / А.В. Панин, Л.П. Худякова // Репродуктивная биология редких и исчезающих растений: тез. докл. Всерос. науч. конф. — Сыктывкар: Коми науч. центр УрО РАН, 1999. — С. 124.
- Сивцева Е.А. Модульная организация и модели побегообразования Pulsatilla flavescens (Zucc.) Juz и Pulsatilla multifida (G. Printz.) Juz. в Якутии / Е.А. Сивцева // Вестн. СВФУ им. М.К. Аммосова. — 2019. — № 2(70). — С. 19–25.
- Цибанова Н.А. Жизненный цикл и возрастная структура ценопопуляций Pulsatilla patens (L.) Mill. (Ranunculaceae) в Северной степи / Н.А. Цибанова // Бот. журн. — 1976. — Т. 61, № 9. — С. 1272–1276.
- Серебряков И.Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных / И.Г. Серебряков. — М.: Высш. шк., 1962. — 380 с.
- Заугольнова Л.Б. Ценопопуляция растений (очерки популяционной биологии) / Л.Б. Заугольнова, Л.А. Жукова, А.С. Комаров, О.В. Смирнова. — М.: Наука, 1988. — 184 с.
- Шаповалова А.А. Динамика возрастной структуры прострела лугового (Pulsatilla pratensis Mill.) в Балашовском районе Саратовской области / А.А. Шаповалова // Экосистемы. — 2016. — Вып. 8. — С. 59–62.
- Заугольнова Л.Б. Типы возрастных спектров нормальных ценопопуляций растений / Л.Б. Заугольникова // Ценопо- пуляции растений (основные понятия и структура). — М.: Наука, 1976. — С. 81–92.