АННОТАЦИЯ
Представлены методологические основы регуляции тканевой гипоксии при неотложных состояниях в медицине поэтапно:вначале указаны механимы биологического окисления и роль тканевого дыхания в митохандриальной дыхательной цепи клеток. Высвобождаемая в процессе метаболизма веществ энергия формируется расшиплением питатетльных субстратов (углеводов, белков, жиров) на трех фазах с дальнейшим переходом в цикл Кребса.
Клинико-экспериментальными исследованияминаучно-обонованы методологические основы регуляции всех форм тканевой гипоксии при неотложных состояниях в медицине.
Ключевые слова: методология, тканевая гипоксия, дыхательная цепь, регуляция, неотложные состояния.
Одной из ведущих проблем современной медицинской науки и практики является гипоксия, так как нарушение кислородного баланса организма человека наблюдается практически при любых патологических состояниях. Недостаточное снабжение тканей кислородом сопровождается изменением метаболизма, функционально-структурного состояния практически всех физиологических систем организма. Это губительно воздействует на весь жизненный цикл, и в первую очередь на ЦНС, что является одной из главных причин смерти больных [1,2].
Гипоксия (hypoxia; греч. hypo +лат, oxy [genium] кислород; син: кислородная недостаточность, кислородное голодание) - состояние, возникающее при недостаточном снабжении тканей организма кислородом или нарушение его утилизации клетками тканей в процессе биологического окисления и/или сопряжения окисления с фосфорилирированием. Поэтому, термин гипоксия обычно трактуют не только как пониженное содержание кислорода в тканях, но и как нарушение процесса утилизации уже доставленного к тканям кислорода и субстратов в необходимом количестве [1,3,4].
К сожалению, при хорошей изученности внешнего дыхания, механизмы тканевого дыхания, то есть процессы утилизации кислорода тканями, в научной литературе мало освещены.Вместе с тем, при неотложных состояниях в условиях отделения интенсивной терапии и реанимации, у больных наблюдается выраженная гипоксия и эффективность кислородной терапии снижается при оксигенотерапии в кислородной палатке и даже при ГБО. Учитывая сказанное, при всех патологических условиях следует оценить эффективность биологического окисления, являющегося основным источником богатых энергией фосфорных соеденинений, необходимых для соответствующей активности функций и обновления структур органов и тканей [2,4]. Обычно окислительные процессы в организме используются клеткой для создания и пополнения ресурсов энергии и для биосинтеза многих соединений (стеринов, простагландинов, нейромедиаторов и др.) и в процессе превращения крупных органических молекул в более простые и, наконец, для образования конечных продуктов обмена веществ и энергии, подлежащих выделению из организма [4,5]. Это диктует необходимость детального изучения механизмов биологического окисления и формирования энергетического обмена.
Между тем, процессы биологического окисления не простое присоединение кислорода к окисляемым субстратам (как некоторые неправильно понимают). Согласно молекулярным механизмам дыхания, биологическое окисление- это сложный ферментативный процесс, протекающий в организмах животных и растений, а также в микроорганизмах. При этом, процесс окисления любого субстрата сопровождается переносом электронов и протонов (т.е. в целом - атомов водорода - дегидрирования) от органических веществ на кислород, то есть от соединения - донора к соединению акцептору. При аэробном окислении конечным акцептором
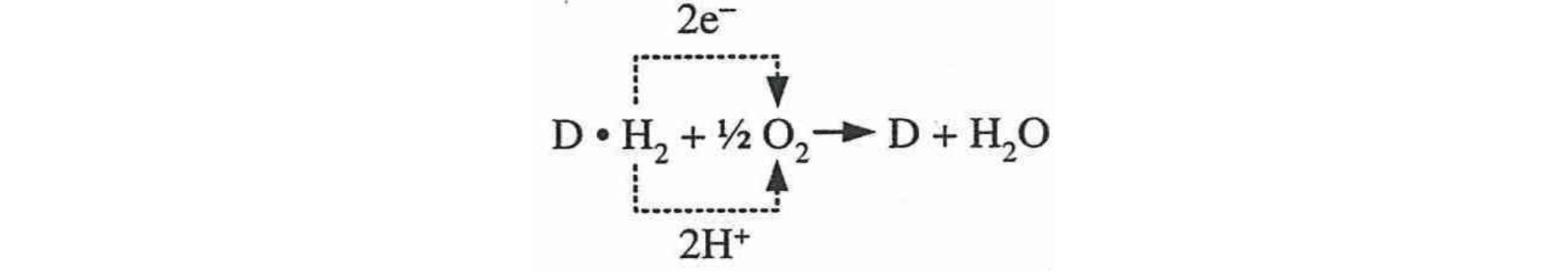
электронов в цепи последовательных окислительно-восстановительных реакций является кислород [1,5,6]:
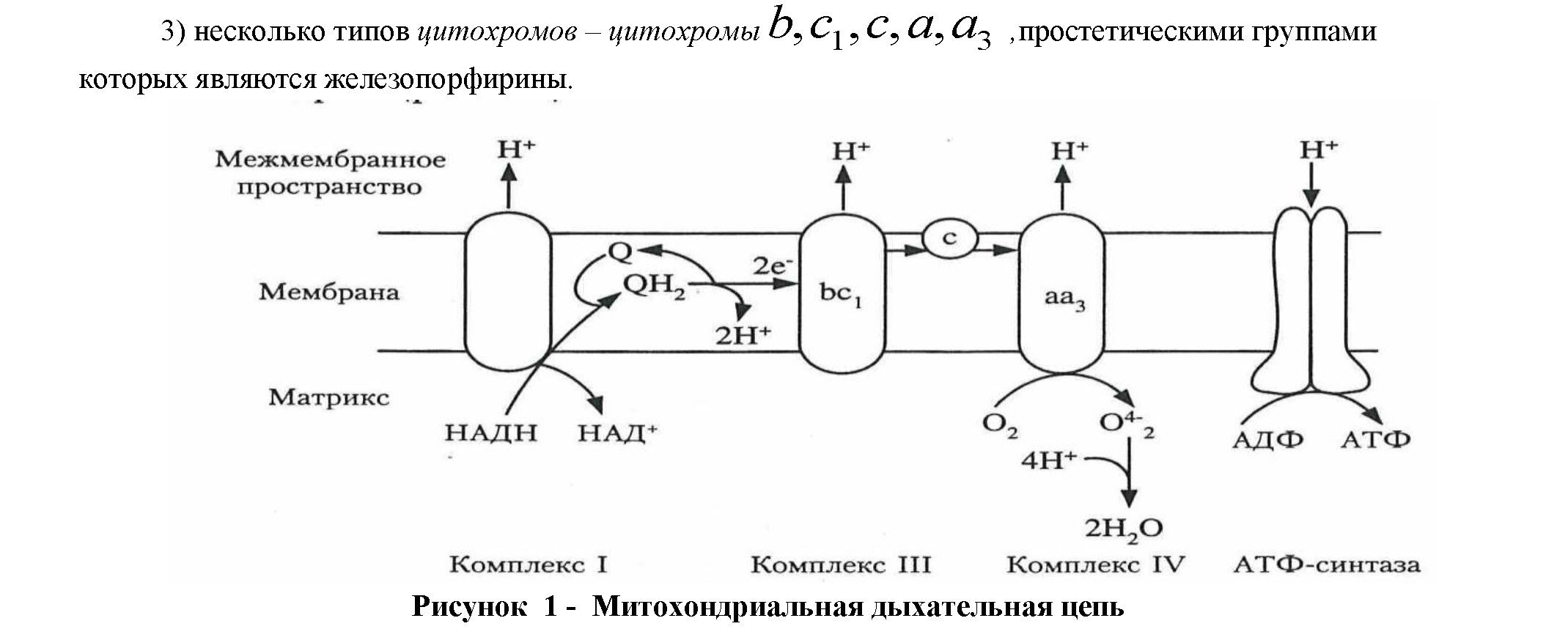
Этот процесс включает много этапов: в нем участвует ряд каскадной системы промежуточных переносчиков, образующих цепь переноса электронов и протонов. Такие реакции катализируютдегидрогеназы - оксидазы (их более 150). При этом в переносе электронов от органических субстратов к кислороду, участвуют дыхательные ферменты (д.ф.), которые являются важнейшим звеном процесса преобразования энергии в дыхательной цепи.В процессе окисления от субстратов и возникающих из них промежуточных продуктов, отнимается водород и, с помощью ферментов, передается в кислород. Эти ферменты, являющиеся катализаторами обменных процессов и, прежде всего, осуществляющих дегидрирование и фосфорилирование, находятся в митохондриях [6,7].
В зависимости от химической природыкофакторов, д.ф. или их простетических групп, подразделяются на три основных класса(рис.1):
1) пиридин-зависимые дегидрогеназы,коферментами которых являются НАД или НАДФ;
2)флавиновыедегидрогеназы, содержащие в качестве простетических групп флавинмононуклеотид (ФМН) или ФАД;
Роль тканевого дыхания в живых клетках чрезвычайно велика, т.к. именно благодаря ему в клетке создается запас основной части энергии, заключенной прежде всего в сложных органических молекулах различной структуры и трансформированной в легко утилизируемой свободной энергии фосфатной связи молекулы АТФ.Тканевая гипоксия наблюдается при блокировании или инактивации, а также разрушения тканевых ферментов биологического окисления и повреждения мембран клеток дыхательной цепи, катализирующих окислительные процессы (оксидазы) в тканях, в результате чего клетки не могут использовать доставленный кислород [2,5,7,11].
В то же время, в качестве источника энергии клетка использует различные субстраты органических веществ пищи.
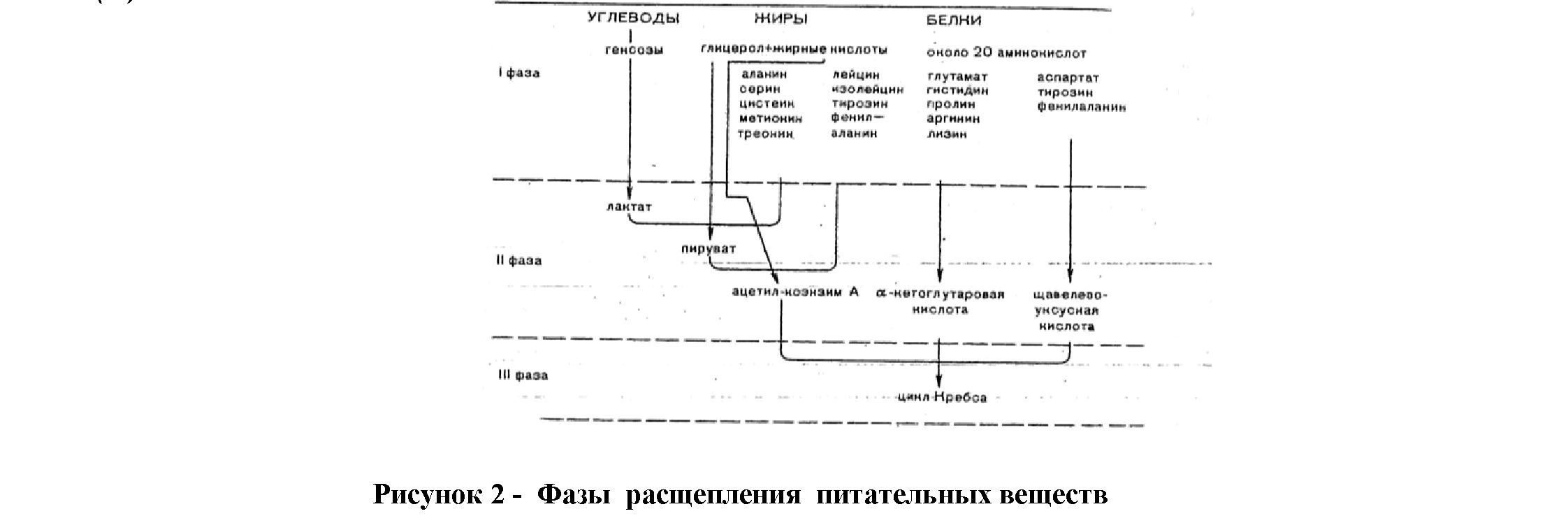
По отношению к высвобождаемой энергиипри метаболизме веществ процессы обмена подразделяются на три фазы(рис.2).
В результате этих процессов из 25-30 веществ образуется, кроме СО2 и Н2О, только три конечных продукта: а-кетоглютаровая, щавелевоуксусная кислота и уксусная кислота в виде ацетил-коэнзима А (ацетил-КоА). Количественно при этом преобладает ацетил-коэнзим А (ацетил-КоА) с дальнейшим их расщеплением в цикле Кребса(рис. 3).
Цикл Кребса является общим конечным путем расщепления как углеводов, так и белков, и жиров. Это как бы узловой пункт в обмене, где сходятся превращения различных структур и возможен взаимопереход синтетических реакций [8,9,10].
Таким образом, в качестве источника энергии клетка использует различные субстраты органического происхождения, которые в ходе окислительного процесса проходят ступенчатые изменения, результатом которого является образование воды и двуокиси углерода.
Следовательно, центр окислительного процесса представляет цикл Кребса, где происходит аэробный гликолиз, распад жирных кислот и обмен аминокислот. Ключевую позицию в обмене занимает ацетил-коэнзим (ацетил КоА), в котором сходятся все пути обмена, прежде чем они перейдут в цикл Кребса. В конечном итоге, состояние интенсивности тканевого дыхания и окислительного фосфолирования определяется продукцией показателей макроэргических
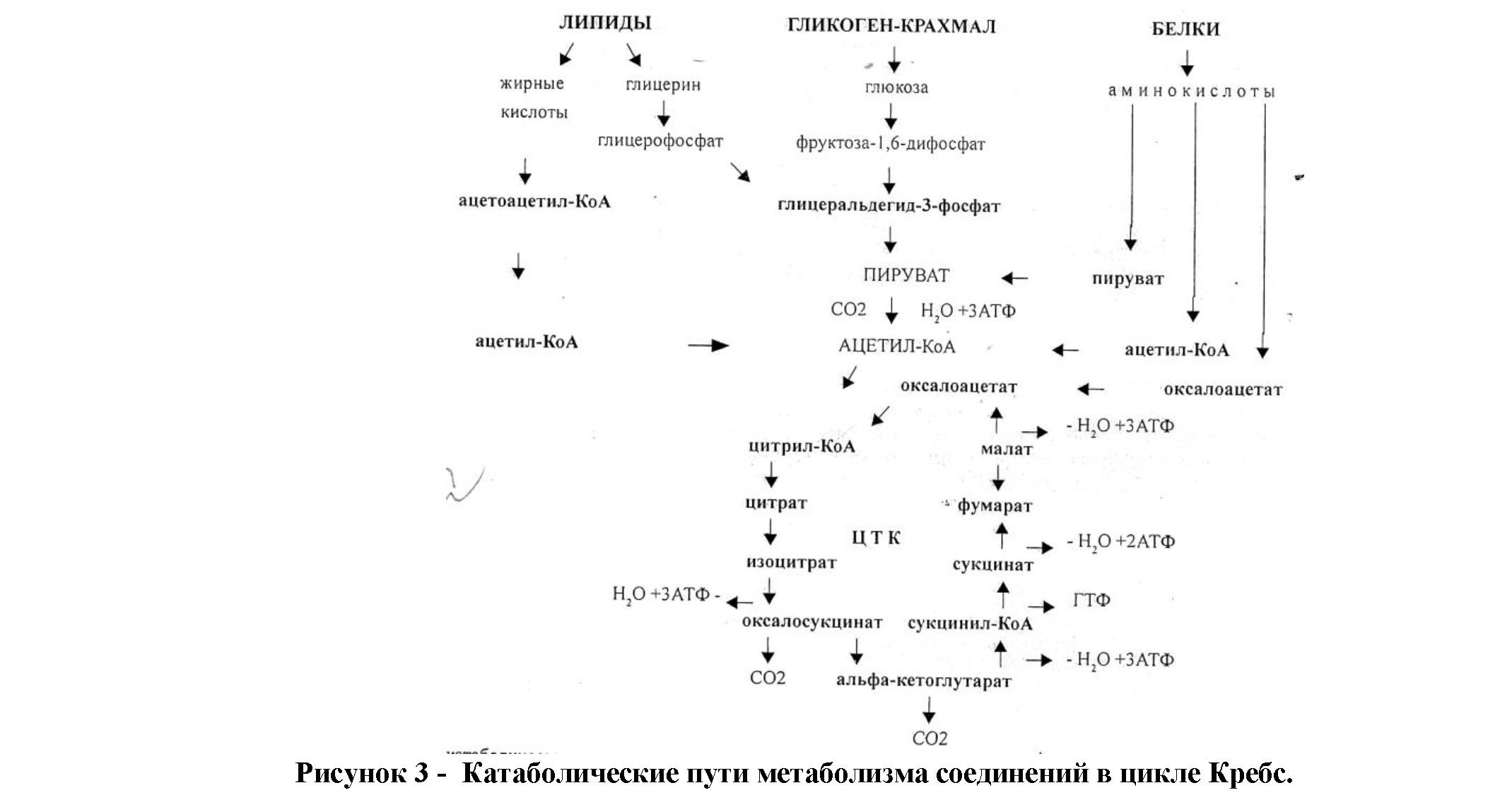
фосфорных соединений (АТФ, ЦТФ, ГТФ, АДФ, АМФ и др.) с вычислением энергетического заряда клеток (1,2,5,7).
Поэтому весь этот сложный цикл интенсивности тканевого дыхания может быть оценен изучением показателей основных участников этапов превращения энергетического обмена.
Развитие этого положения легло в основу проведенных наших клинико-экспериментальных исследований[12,13,14]. Результатами проведенных исследований у больных с респираторной патологией научно обоснован и апробирован новый системный подход в регуляции тканевой гипоксии, заключающийся в использовании естественных переносчиков электронов и протонов субстрат, направленный на улучшение тканевого дыхания путём шунтирования инактивированных участков дыхательной цепи,под защитой энергетического аппарата клеток через стабилизацию биологических мембран на фоне витамино-энергетического комплексав следующей
последовательности:
- НАД или никотинамид;
- ФАД (флавинат) или рибофлавин-мононуклеотид;
- убихинон (коэнзимО);
- цитохром С.
Это открывает новые перспективы в области применения эффективных фармакопрофилактических и терапевтических средств в регуляции всех форм тканевой гипоксии в медицинской практике.
СПИСОК ЛИТЕРАТУРЫ
- Окисление биологическое//Большая медицинская энциклопедия, издание 3, том 17, - Москва. Издательство «Советская энциклопедия», -1981. -С.250-255.
- Дудко В.А., Соколов А.А. Моделированная гипоксия в клинической практике. //В.А. Дудко, А.А. Соколов. - Томск: STT, -2000, -352с.
- VanLiereEJ., StickneyJ.C. /Ван Лир Э. и Стикней К. Гипоксия.пер. с англ., М.-1967.
- Колчинская А.З. О классификации гипоксических состояний //А.З. Колчинская.Патол. физиология и эксперимент.терапия,- 1981, № 4, -С.3-10.
- Николаев А.Я. Тканевое дыхание, дыхательная цепь //А.Я. Николаев. Биологическая химия. - Москва - 2007. - С. 225-236
- Скулачёв В.П. Транспорт энергии, метаболитов, кислорода и электронов вдоль биологических мембран // В.П. Скулачёв. Успехи соврем.биол.1979.,- т.38.№2(5).-С.163—180.
- Лукъянова Л.Д. Биоэнергетические механизмы формирования гипоксических состояний и подходы к их фармакологической коррекции // Л.Д. Лукъянова.Фармакологическая коррекция гипоксических состояний. - Москва, 1989. - С. 11-44.
- Мак Мюррей У. Обмен веществ у человека. // У. Мак Мюррей - М.Мир.1980,-368с.
- Шарманов Т.Ш.,Мухамеджанов Э.К. К проблеме взамосвязи между обменом белков, жиров и углеводов // Т.Ш. Шарманов, Э.К. Мухамеджанов. Вопросы питания -1982, №4, - С. 10-16
- Тапбергенов Салават Медицинская биохимия.// Салават Тапбергенов. - Астана, 2001, Фолиант,-292 с.
- Ванников Л.Л. Антигипоксантное действие ПСН // Л.Л. Ванников. Тканевая гипоксия и ее коррекция.-Новосибирск.:Наука, - 1981.-С.4-27.
- Мусаев Ю.М. Эффективность коферментов и мембраностабилизирующих средств на фоне витамино-энергетического комплекса при интенсивной терапии пневмонии новорожденных и детейраннего возраста// Ю.М. Мусаев.ХІІ Всесоюзный съезд детских врачей. - Москва, 1988. - С.257-258.
- Мусаев Ю.М. Клинико-молекулярные основы гипоксического синдрома и новые подходы к его метаболической регуляции при острой осложненной пневмонии у детей раннего возраста: // Ю.М. Мусаев.Дис. ... доктора мед. наук, -Ташкент. 1993, - 343с.
- Мусаев Ю.М. Современные представления о гипоксическом синдроме у детей с острой пневмонией // Ю.М. Мусаев.Вестник Казахского национального медицинского университета - 2006 - №3 - С.86-90.