В статье проведен литературный обзор по проблемам влияния на показатели нейрогуморальной регуляции стрессовых факторов. Показано, что имеющиеся определенные механизмы устойчивости организма к стрессовым факторам связаны с особенностями деятельности нервной системы отдельных особей. В зависимости от типа высшей нервной деятельности раскрыты нейрогуморальные и молекулярные механизмы стрессов. Представлены данные о роли гормонов и нейромедиаторов центральной нервной системы. По литературным источникам дан анализ исследованиям о формировании защитных механизмов организмов при действии физических факторов.
Термин «стресс» приобрел особое значение в науке о высшей нервной деятельности (ВНД) [1-7].
Известно, что научное использование термина «стресс» ввел канадский ученый Г.Селье и обозначил этим термином общие, неспецифические изменения в системе гипофиз-надпочечники, возникающие на любые «необычные» (стрессорные) воздействия. При этом речь шла о сильных болевых, токсических и других, в том числе психических воздействиях, а критерием «неспецифичности» являлась главным образом гуморальная система.
Вторым важным моментом концепции Г.Селье является функциональная однородность, стадийность и неспецифичность стресса. Любой стресс проходит в своем развитии 3 стадии: тревоги, резистентности, истощения.
Третий момент заключается в адаптационном значении стресса.
Современными исследованиями [5] при моделировании на животных некоторых психоневрологических заболеваний методом условных рефлексов и образного поведения было показано, что информационный невроз, возникающий в условиях неограниченной двигательной активности, вызывающий формирование стресса, происходит вследствие длительного влияния на высшую нервную деятельность триады факторов: большого объема информации, дефицита времени, необходимого для ее обработки, и высокого уровня мотивации поведения.
Данная триада имела значительные индивидуальные различия, которые носили многоплановый характер и отражались на поведенческих вегетативных, соматических, нейрофизиологических, биохимических и других показателях. Она определяла возможность восстановления функций после прекращения воздействия стресс-факторов, а также латентный период формирования донозологических (предпатологических) симптомов и манифестацию патологии, характер нелекарственной и лекарственной коррекции. Поэтому, естественно, что учет усредненных показателей совершенно не отражает истинную природу проявления стресса.
Начиная с 1977 г. К.В.Судаковым и его школой [1-4] обоснованы индивидуально-типологические подходы при изучении эмоционального стресса. При этом процент устойчивости адаптирующихся и предрасположенных к изменению сердечно-сосудистой системы (ССС) у белых крыс был разным в зависимости от генетической линии — наиболее устойчивыми оказались линия Вистар, наименее — Август.
Авторы отмечают, что у всех животных, как у устойчивых, так и у неустойчивых, наблюдались все фазы стресса, открытые Селье, т.е. проявлялись неспецифические его механизмы. Наряду с этим у животных были обнаружены как устойчивость одних функциональных систем, так и неустойчивость других: в однотипных конфликтных ситуациях у одних были специфические нарушения со стороны ССС (АД и др.), у других — со стороны желудочно-кишечного тракта (активация адренер-гической иннервации и др.), у третьих — функции щитовидной и других желез внутренней секреции.
В настоящее время считают, что характер поведения крыс в «открытом поле» является наиболее доступным и легко измеряемым прогностическим критерием устойчивости крыс к стрессу. Показано, что крысы, проявляющие короткий латентный период первого движения и выхода в центр, а также высокую двигательную активность по периферии и, особенно, в центре, относятся к устойчивым.
Крысы, демонстрирующие продолжительный латентный период первого движения и выхода в центр, низкую активность в центре и на периферии и имеющие высокие цифры вегетативного баланса, были предрасположены к эмоциональному стрессу.
При исследовании содержания нейрорецепторов и нейромедиаторов в лимбико-ретикулярных структурах мозга было показано, что у крыс, устойчивых к эмоциональному стрессу по показателям выживаемости, было исходное достоверно высокое содержание норадреналина (НА) в гипоталамусе и низкий уровень дофамина (ДА) в области перешейки и среднего мозга. Поэтому приходят к выводу, что устойчивость к стрессу определяется сложными, генетически обусловленными особенностями нейрохимических процессов в различных структурах мозга. При эмоциональном стрессе в основе центральных механизмов, определяющих устойчивость к стрессу, играет роль специфическая организация молекулярных и нейрохимических свойств нейронов гипоталамо-лимбико-ретикулярных структур мозга.
Индивидуальный подход позволяет направленно изменять исходную нейрохимическую организацию структур мозга при дополнительном введении олигопептидов (например, вещества Р), которые долговременно повышают содержание НА в гипоталамусе [3].
С другой стороны, доказано изменение чувствительности нейронов коры и подкорковых структур мозга к медиаторам и олигопептидам. Этот процесс при стрессе имел сложный интегративный характер, т.е. мозг как интегрированная целая структура в условиях выраженной конфликтной ситуации изменяет свои нейрохимические свойства. Длительное сохранение этой ситуации обусловливает переход этих изменений в устойчивую стационарную форму (патологию).
Считают, что природа нейрохимической интеграции «застойного» возбуждения полихимична — в ее механизмах, наряду с олигопептидами, играют роль и медиаторы, которые действуют на фоне уже измененных нейронов и, по-видимому, глиальных элементов нервной ткани мозга.
Таким образом, на примере эмоционального стресса доказано, что устойчивость к стрессу в значительной степени зависит от индивидуальных особенностей саморегуляции различных функциональных систем в организме, слаженности их иерархического и мультипараметрического взаимодействия по конечным приспособительным результатам их деятельности [4]. При действии физических факторов эти механизмы не изучены.
Индивидуально-типологический анализ различных показателей жизнедеятельности у животных в однотипных конфликтных ситуациях, порождающих стресс, имеет значительные преимущества в плане понимания нейрогуморальных и молекулярных механизмов стресса.
Среди функциональных систем, участвующих в организации приспособительных реакций и сохранении устойчивости гомеостаза, важная роль принадлежит нейрогуморальной системе. Из общей массы метаболитов, циркулирующих во внутренней среде и принимающих участие в регуляции и корреляции процессов жизнедеятельности организма, ведущее место занимают биологически активные вещества. Известно, что биологически активные вещества (БАВ) образуются при декарбоксили-ровании аминокислот (АМК) и обладают чрезвычайным и мощным фармакологическим действием.
К ним относятся гистамин, серотонин и катехоламины. Нарушения обмена их может быть ведущим и играть роль «пускового механизма» при срыве адаптации организма [6-10].
Диапазон их влияния на функции организма широк и многообразен — тканевые гормоны, медиаторы, регуляторы внутриклеточных, тканевых, органных превращений [11-15].
Катехоламины (КА) вырабатываются в надпочечниках (в мозговом его слое), из аминокислоты тирозина (паратоксифенилаланина); образовано из незаменимой АМК-фенилаланина.

Окисление тирозина обусловлено внедрением атома кислорода в циклическую часть молекулы с образованием ДОФА (диоксифенилаланина) — Д. Затем ДОФА подвергается декарбоксилированию и образуется ДОФамин (окситирамин) — ДА, у которого окисляется боковая цепь и образуется но-радреналин (НА). При взаимодействии с активной формой S-аденозилметионина в боковой цепи но-радреналина метилируется группа NH2 и образуется адреналин (А).
Физиологические трофические эффекты внесинаптического действия КА выявлены при воспроизведении феномена Орбели-Гинецинского. Оказалось, что при внесинаптическом контакте с мышечными клетками А и НА вызывают такой же адаптационно-трофический эффект, как и раздражение симпатических волокон. Внесинаптическое адаптационно-трофическое действие КА, особенно А, проявляется во время динамической работы, при которой в кровь выделяется повышенное количество А из мозгового вещества надпочечников, НА при этом выделяется менее интенсивно. К адаптационно-трофическим эффектам в данном случае относят хроно- и инотропное действие КА на сердце, гликогенолиз, липолиз, активацию гипофизарно-адреналовой системы (повышение энергического и других видов обмена), восходящей ретикулярной формации мозга, усиление кровоснабжения большинства органов и как результат — повышение тонуса кровеносных сосудов, умственное возбуждение, состояние тревоги или уверенности и агрессивности (в зависимости от типа ВНД).
Часть перечисленных адаптационно-трофических эффектов действия КА отчетливо проявляется при срочном несильном и непродолжительном действии холода на неадаптированные к нему организм, а также при срочном сильном действии холода на адаптированные к низким температурам организмы. В архитектуру адаптационных изменений при действии холода включаются, вслед за активацией САС, гиперфункция и гипертрофия коры надпочечников, щитовидной железы, индуцированный КА и гормонами указанных желез, приспособительный рост массы и функциональной мощности системы митохондрий в мышцах и бурой жировой ткани, увеличение массы и физиологической активности всех звеньев систем, ответственных за транспорт к митохондриям кислорода и субстратов окисления, а тем самым за усиленное повышение теплопродукции и резкое снижение теплоотдачи.
Нервно-психическая нагрузка сопровождается существенной стимуляцией САС, резко выраженной эрготропной реакцией при сравнительно умеренном уровне ряда перечисленных адаптационно-трофических эффектов, являющихся следствием внесинаптического действия КА.
Адаптационно-трофическое влияние КА четко выявляется в опытах на животных с внутривенным введением их в физиологических дозах, а также с электрофоретической аппликацией этих веществ на клетки нервной и других тканей, при перфузии изолированных органов растворами, содержащими КА [16, 17] и особенно тех, функция которых выражается биосинтезом и секрецией нужных для организма веществ и выведением ненужных метаболитов. Биосинтез веществ железами внешней секреции и клетками других органов под влиянием КА и других медиаторов по своей сути является процессом трофическим, который без очищения клеток от конечных и промежуточных продуктов метаболизма осуществиться не может. Это относится и к нейронам всех уровней нервной системы, так как любому нейрону свойственна нейросекреция, проявляющаяся аксоплазматическим током веществ, выведением продуктов биосинтеза в жидкие среды организма и в клетки через синапсы. Это относится и к клеткам эпифиза, инкреторная функция которого у млекопитающих регулируется симпатическим нервом. Отмечено значение КА в генезе ретардации роста физиологически незрелых крыс в постнатальном периоде, что подтверждается более низкой скоростью роста тела и мозга и нарастания содержания в них КА. Интересно, что деградация энкефалинов, обладающих трофической функцией, ингибируется КА путем инактивации аминопептидаз [18].
В сложных механизмах нервнотрофического обеспечения функции организма существенная роль принадлежит прямому внесинаптическому действию на клетки тканей и органов циркулирую
щему в жидких средах тканевого гормона серотонина (СТ), который образуется как в нервной, так и в других тканях организма [18-20]. Особенно много его в тромбоцитах крови и в ЖКТ.
СТ образуется из АМК-триптофана окислением его при участии триптофангидроксилазы.

Активность СТ зависит от наличия в его молекуле оксигруппы в положении 5 и свободной аминогруппы. Окисление СТ моноаминооксидазой (МАО) дезактивирует гормон.
СТ является важным тканевым гормоном и нейромедиатором в ЦНС, поддерживающим нормальную психическую деятельность. Действие СТ на нервную систему осуществляется в тесном взаимодействии с А и зависит от состояния нервной системы. Серотонин жидких сред организма, наряду с функцией межклеточной передачи сигналов, обладает модулирующим влиянием на внеклеточный метаболизм веществ [19, 20]. Изменение содержания СТ в крови и тканях отражается на обмене нуклеиновых кислот, белков, липидов, углеводов, на уровне энергетического метаболизма, сопряженности окисления и фосфорилирования, биосинтеза адениновых нуклеотидов. Однако не только прямое участие серотонина в регуляции внутриклеточного метаболизма определяет его участие в регуляции трофики клеток. Серотонину, так же как и веществу Р, другим полипептидам, простаглан-дину Е, приписывают функцию модулятора синаптической передачи в вегетативной системе. Показано, что СТ потенциирует действие А и НА [19, 18], участвует в передаче возбуждения в ганглиях интрамуральной нервной системы в ЖКТ [21], а также в центральной регуляции вегетативной нервной и эндокринной систем, активируя или модулируя деятельность нейронов ядер переднего гипоталамуса, его аденогипофизотропной, эрго- и трофотропной зон. Следовательно, СТ и непосредственно, и опосредованно может оказывать адаптационно-трофическое влияние практически на все органы и ткани.
Как и КА, СТ, в зависимости от локализации его введения в надгипоталамические и гипотала-мические образования, может активировать или угнетать систему гипоталамус — аденогипофиз — периферические железы внутренней секреции.
Непосредственное влияние А (8 нг) и СТ (40 нг) на нейроны нервных структур, окружающих желудочки мозга, при субокципитальном 10-кратном введении этих веществ в 0,5 мл физиологического раствора в течение 30 суток (одно введение каждые 3 дня) предотвращало развитие язв, уменьшало частоту, скорость возникновения и размеры облысения на левой конечности кроликов, у которых перерезали левый седалищный нерв [18].
Имеются данные о негативных трофических эффектах от больших количеств эндо- и экзогенного СТ. Сообщается об ульцерогенном его действии при повышенных концентрациях в крови. Отмечены фазные изменения содержания СТ в отделах желудка и головного мозга при нейрогенной дистрофии ЖКТ.
Введение СТ животным приводило к изменению функциональной активности всех желез внутренней секреции, что было обусловлено не только прямым его действием на эти органы, но и опосредованным влиянием через гипоталамо-нейрогипофизарную, гипоталамо-аденогипофизарную и вегетативную нервную системы, а также через изменение чувствительности эндокринных органов к специфическим гуморальным раздражителям, уровень которой определяется трофическим состоянием этих органов [18].
Гистамин (ГН) — нейромедиатор, тканевой гормон, общепризнанный периферический регулятор тонуса и проницаемости мембран и капилляров, объема циркулирующей крови, преимущественно оказывающий влияние на проницаемость эндотелия венул, наиболее выраженное в коже и в мышцах [22-24]. (Гистамин синтезируется из аминокислоты гистидина путем его декарбоксилирования при участии кофермента фосфопиридоксаля).
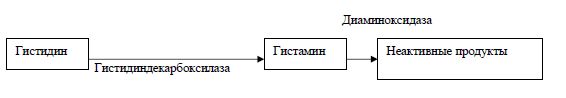
Встречается гистамин (ГН) в связанном состоянии с гетерополисахаридом (гепарином), ацетатом и другими веществами.
Предполагают, что ГН может находиться в виде «клешневидного» комплекса с тяжелыми металлами, когда металл соединяет две молекулы между собой за счет аминных групп. Избыточное накопление ГН вызывает шок. Естественными регуляторами ГН в тканях являются КА (А и НА), аце-тилхолин, гормон роста. При изучении возрастных изменений гистамина в тканях крыс показано его депрессирующее действие на клеточный геном через активацию протеинкиназы, т.е. гистамин проявлял себя как модулятор роста.
За последние годы накопились новые сведения о специфических особенностях обмена гистами-на у животных и человека. Особенности действия гистамина выявлены при изучении динамической системы субстрат-фермент (гистамин-диаминоксидаза-гистаминопексия), а также во взаимодействии с другими биологически активными веществами, например, с серотонином. Доказано наличие специфических рецепторов к гистамину (Н1 — в гладких мышцах и больших кровеносных сосудах; Н2 — в различных тканях, в том числе в гладких мышцах). При стимуляции Н1-рецепторов вызывается сокращение, или сужение, сосудов. При стимуляции Н2-рецепторов наблюдается расслабление, расширение сосудов. Считают, что Н2-рецепторы могут играть значительную роль в развитии гипереми-ческой реакции, вызванной физической нагрузкой.
Гистамин (ГН) стимулирует гипофизарно-надпочечниковую систему, вызывая процессы возбуждения в различных структурах мозга (гипоталамусе, обонятельном мозге, мозжечке). Выявлено влияние гистамина на образование и секрецию ряда гормонов — АКТГ, гормонов гипофиза и коры надпочечников, которые, в свою очередь, по «принципу обратной связи» воздействуют на обмен гис-тамина.
Источником гистамина в организме человека и животных являются различные клетки: тучные клетки соединительной ткани, базофилы, лимфоциты, эозинофилы, тромбоциты и др. Гистамин депонируется в них, образуя лабильные комплексы с гепарином, АТФ и РНК. В ткани, где тучные клетки отсутствуют, он может быть связан с гликофосфатом [25]. Совместное введение малых доз серо-тонина и гистамина значительно повышало устойчивость животных к гипоксии, что свидетельствовало об участии этих аминов в механизмах приспособительных реакций. В механизме защитного действия важная роль отводится их способности ингибировать перекисное окисление липидов в биомембранах [23].
Считают, что в организме животных и человека ГН может быть образован из нескольких источников:
1) поступать (около 5 %) с пищей (молоко содержит 0,5 мкг/г и т.д.);
2) образоваться из белков пищи при декарбоксилировании гистидина бактериями кишечной флоры группы Coli (т.н. «экзогенный» гистамин);
3) образоваться при внутриклеточном декарбоксилировании гистидина ферментом — гистидин-декарбоксилазой (т.н. «эндогенный» гистамин) в ряде паренхиматозных органов и в мозге. Этот процесс протекает с разной скоростью. Например, в печени эмбриона крысы активность фермента в 3000-5000 раз больше, чем в печени матери. Показано значительное ускорение синтеза в селезенке эмбрионов, т. е. в органах с высокой гемопоэтической активностью, что указывает на связь гистами-нобразующей активности с митозами клеток в период эмбрионального развития.
Биосинтез ГН в тканях млекопитающих осуществляется ферментативным декарбоксилировани-ем АМК — гистидина при участии гистидиндекарбоксилазы (ГД), как специфической, так и содержащейся в микрофлоре кишечника. ГД является фосфопиридоксалем протеидов, поэтому для его активности необходим витамин В6.
Синтез ГН отличается своей простотой, так как для серотонина требуется еще окисление ин-дольного кольца, а для норадреналина — метилирование боковой цепи.
Инактивация свободного, биологически активного гистамина происходит под действием диами-ноксидазы (ДО), обнаруженной в микросомах и митохондриях печени крыс. Показано наличие в активном центре ДО (гистаминазы) пиридоксальфосфата и меди.
Основным путем обмена гистамина во многих тканях и органах, и особенно интенсивно в гипоталамусе, является метилирование азота в имидазольном кольце гистамина с последующим окислением метилгистамина в присутствии моноаминооксидазы (МАО).
И.Л.Вайсфельд и Г.Н.Кассиль [22] при проведении функциональных нагрузок отмечают быстрое восстановление ГН до исходных уровней и считают, что для регуляторных систем, синтезирующих и инактивирующих ГН, характерны высокая надежность в обеспечении постоянства содержания ГН в организме.
По данным ряда авторов, ГН в организме находится: 1) в свободном физиологически активном состоянии, но количество его незначительно; 2) в лабильносвязанном состоянии, легко переходящем при воздействии разных факторов (стрессе, физическом воздействии, патологических процессах и др.) в свободный; 3) прочносвязанный, который освобождается только в случае механического разрушения тканей при кислотном гидролизе или действии высокой температуры.
В виду того, что ГН освобождается из тканей под действием протеолитических и гемолитических ферментов, считают, что ГН связан пептидными связями со свободными карбоксильными группами аминокислот в составе белков.
Однако освобождение ГН из тканей при неферментативных воздействиях дало основание предположить, что ГН в тканях может соединяться через солеобразные ионные связи с кислотой (типа гепарина). В тучных клетках обнаружен тройной комплекс «гепарин-цинк-гистамин». Микроэлементом в данном случае может быть и свинец. В тканях без тучных клеток ГН может быть связан с РНК, дифосфопиридиннуклеотидом, АТФ.
Белки могут связывать ГН в нейтральной среде и отдавать его при действии более сильных ка-тионно-щелочных ионов буфера. Разведенная в 20 раз сыворотка крови здоровых людей снижала биологическую активность ГН in vitro на 30 % — этот эффект получил название «гистаминопексиче-ского эффекта» (ГПЭ). Установлено его снижение у больных аллергией и другими заболеваниями. Этот эффект характерен и для крови крыс и относится к гамма-глобулинам или другим белкам с большим молекулярным весом, чем у иммунноглобулинов G, и гликопротеидам. Показано, что избыток К+ тормозит связывание ГН, а избыток Са2+ усиливает. Удаление надпочечников и гипофиза у крыс приводит к полной потере инактивации ГН сывороткой.
Большое число исследований посвящено изучению ГН в ЦНС и периферических отделах нервной системы. Распределение и количественное его содержание имеет важное значение для понимания роли ГН в регуляции функции.
Распределение ГН в периферических нервах выявило интересные факты:
1) количество его ступенчато увеличивается от спинного мозга к спинным корешкам и далее к постганглионарным симпатическим нервам;
2) в чувствительных нервах ГН больше, чем в двигательных. Считают, что в периферических нервах местом образования гистамина являются не только тучные клетки, но и сама нервная ткань.
В определенных структурах ЦНС показано наличие ГН и ферментных систем, участвующих в его биосинтезе и распаде.
Уровни эндогенного гистамина в мозге по разным методикам определяются от 0,05 до 6,8 мкг/г ткани. Была доказана связь гистамина с ростом отростков нервных клеток (заметное увеличение в гипоталамусе между 40 и 44 днем развития плода морских свинок). К рождению уровень гистамина в мозге достигает его уровня у здоровых морских свинок, так же как и активность метилтрансферазы. Высокое содержание не только ГН, но и всех аминов (АХ, НА, СТ) обнаружено в гипоталамической области.
Неравномерное распределение ГН в различных отделах мозга зависит и варьирует от вида животных, возраста, линии в пределах одного вида. Изучение субклеточного распределения ГН показало, что наибольшее содержание его обнаруживается в микросомах и коррелирует со значительным числом нервных окончаний и синаптических везикул в микросомальной фракции.
Содержание ГН в митохондриях мозга у крыс выше, чем СТ и АХ, но близко к распределению НА и ДА. Подтверждением этих данных являются публикации о локализации ГН, гистидиндекарбок-силазы (ГД) и гистаминметилтрансферазы (ГМТФ) в микросомальной и митохондриальной фракциях из гомогената ткани гипоталамуса.
Установлена химическая структура содержащихся в мозге комплексов гистамина. Так, внутри микросом они представлены комплексами ГН + сульфомонополисахариды, АТФ, нуклеиновыми кислотами и дифосфопиридиннуклеотидом. Это указывает на связь ГН с энергетическими процессами, протекающими в определенных внутриклеточных структурах нервной ткани.
Доказано активное участие гистамина в регуляции функциональной активности разных нервных структур — синапсов, рецепторов, нервных проводников, т. е. в процессах синаптической передачи и рецепции.
Исследования последних лет значительно расширили и углубили представления о Н1- и Н2-гистаминовых рецепторах. Открытие гистаминергических нервов и специфической системы нейро
нов в головном мозге позволяет рассматривать гистамин как медиатор нервных импульсов в ЦНС. Предполагают, что Н1-рецепторы опосредуют замедление атриовентикулярного проведения, а Н2 участвуют в медиации при увеличении частоты сокращения сердца.
Перфузия сердца животных гистамином вызывала положительное инотропное (снижение возбудимости сердечной мышцы) действие, повышая проницаемость мембран для ионов Са++ и усиления образования цАМН, принимающего участие в формировании каналов медленного переноса Са++.
Гистамин резко нарушает сосудисто-соединительно-тканную проницаемость, усиливая процессы деполяризации в основном веществе соединительной ткани, накопление мукополисахаридов в пе-риваскулярных зонах стромы органов и фибриноидное набухание сосудов.
Повышение проницаемости гисто-гемотических барьеров при лучевых поражениях при действии рентгеновских лучей связывают с накоплением гистамина во внутренней среде.
Считают, что система «гистамин-гистидиндекарбоксилаза» является «внутренним механизмом» регуляции микроциркуляции через следующие механизмы: гистамин синтезируется непрерывно внутри мелких кровеносных сосудов; гистамин действует главным образом на специфические рецепторы; скорость синтеза гистамина зависит от условий окружающей среды и подчиняется механизмам адаптации.
Физическая нагрузка, холод, введение катехоламинов, воспаление, злокачественные опухоли и другие вызывают увеличение активности гистидиндекарбоксилазы и тем самым оказывают влияние на синтез гистамина.
Считают, что введенный парэнтерально гистамин может играть роль универсального пускового механизма — неспецифического стресс-раздражителя. Гистамин в малых дозах является эффективным стимулятором гипофизарно-надпочечниковой системы.
Следовательно, гистамин возбуждает различные структуры мозга и является звеном в сложной цепи реакций, приводящих к секреции АКТГ (адрено-кортикотропного гормона). Доказано наличие значительного количества гистамина в нервных окончаниях и синаптических везикулах коры мозга.
Таким образом, гистамин оказывает стимулирующее влияние на нейроэндокринную регуляцию, осуществляемую промежуточно-гипофизарными образованиями. С другой стороны, доказано осуществление нейрогуморального контроля над обменными процессами гистамина гормонами гипофиза и особенно коры надпочечников на активность гистаминдекарбоксилазы, диаминоксидазы в крови и органах (печень, легкие, почки, мозг), обеспечивающими относительно постоянный уровень гиста-мина в организме.
Опыты с внутривенным введением центрально-действующих веществ (подавляющих синтез и распад, истощающих запасы адреналина и др.) дают основание предполагать, что содержание клеточного гистамина регулируется общими физиологическими механизмами, являющимися регуляторами и других гомеостатических реакций в биологическим организме.
Считается, что гистамин является основным биохимическим медиатором иммунологических реакций. Имеются сведения о корреляции изменения титра противотканевых аутоантител к сосудистым, нервным, хрящевым и костным антигенам и содержания гистамина в крови при вибрационной болезни [26]. Авторы приходят к выводу, что одним из патогенетических механизмов при этом является влияние гистамина на иммунную систему.
Увеличение гистамина коррелировало с увеличением содержания В-розеткообразующих лимфоцитов (В-РОЛ), что может приводить к выработке специфических противотканевых антител, а в механизмах дефицита Т-розекткообразующих лимфоцитов (Т-РОЛ) имеет значение увеличение уровня простагландинов группы ПГФ-2а с последующим ингибированием реакции бластотрансформации лимфоцитов. Выявленные изменения соответствовали тяжести заболевания. По-видимому, гистамин играет определенную роль в механизмах адаптации иммунной системы [27] и возникновении вторичных иммунодефицитов в процессе хронического эколого-профессионального перенапряжения, установленного А.А.Новицким и соавт. [28] и другими исследователями.
Таким образом, приведенные данные подчеркивают многообразие путей, регулирующих содержание КА, СТ и ГН в различных системах организма и обосновывают их участие в стресс-реакциях животных и человека. Однако нами не найдено исследований, касающихся их роли в формировании защитных механизмов при действии физических факторов (мышечной деятельности) у тренированных и нетренированных животных, в зависимости от длительности воздействия и типа ВНД.
Список литературы
1 Судаков К.В. Корково-подкорковые взаимоотношения в условиях острого эмоционального стресса // Журн.высш.нерв.деят. — 1977. — Т. XXVII. — Вып. 2.
2 Судаков К.В. Системные механизмы эмоционального стресса. — М., 1981.
3 Судаков К.В. Механизмы устойчивости к эмоциональному стрессу: преимущества индивидуального подхода // Вестник РАМН. — 1998.
4 Судаков К.В. Теория функциональных систем и профилактическая медицина // Вестн. РАМН. — 2004. — № 5.
5 Хананашвшш М.М. Психогенный стресс: теория, эксперимент, практика // Вестник РАН. — 1998. — № 8. — С. 13-15.
6 Кузьмина Л.П., Тарасова Л.А. Биохимический профиль организма: теоретические и практические аспекты изучения и оценки // Журн. мед. труда и промышл. экология. — 2000. — № 7.
7 Antelman S.M., Chiodo L.A. Stress: its effect on interaction among biogenic amines and role in the induction and treatment of diseases // Handbook of Phycho pharmacology. — 1984. — Vol. 18.
8 КалуевА.В. Проблемы изучения стрессорного поведения. — Киев: CSF, 1998. — С. 35-12.
9 Gromowa E.A., Semenowa T.P. Interrelation on individual reactivity, learning capacity and peculiarities of metabolism of monoamines in the brain // III Congress of the Bulgarian Pharmasia, 17-20 May. — 1990. — P. 45-16.
10 Рогов А.В., Пеккер Я.С., МедведевМ.А., Берестнева О.Г. Адаптационные характеристики человека (оценка и прогнозирование). — Томск: Изд-во Томск. ун-та, 1997. — 279 с.
11 Акмаев И.Г. Современные представления о взаимодействиях регулирующих систем: нервной, эндокринной, иммунной // Успехи физиол. наук. — 1986. — Т. 27. — № 1. — С. 3-21.
12 Барабой В.А. Механизмы стресса // Успехи совр. биологии. — 1991. — Т. 3. — Вып. 6. — С. 23-29.
13 Базаревич Г.Я., Богданович У.Я., Волкова И.Н. Медиаторные механизмы регуляции дыхания и их коррекция при экстремальных состояниях. — Л.: Медицина, 1979. — 200 с.
14 Власов В.В. Реакции организма на внешние воздействия: общие закономерности развития и методические проблемы исследования. — Иркутск, 1994.
15 Острый О.Я. Нервная трофика в физиологии и патологии. — М.: Медицина, 1970. — С. 90.
16 АжипаЯ.И. Трофическая функция нервной системы. — М.: Наука, 1990. — 672 с.
17 Ажипа Я.И. Участие нейромедиаторов в эфферентном звене трофического рефлекса // Физиол. чел. — 1992. — Т. 18. — № 6.
18 КурскийМ.Д., БакшеевН.С. Биохимические основы механизма действия серотонина. — Киев: Наук. думка, 1984. — 296 с.
19 НауменкоЕ.В., ПоповаН.К. Серотонин и мелатонин в регуляции эндокринной системы. — Новосибирск, 1975. — 218 с.
20 ГромаковскаяМ.М. Нейрогуморальные механизмы регуляции мышечной деятельности. — М.: Наука, 1965. — 234 с.
21 Вайсфельд И.Л., Кассиль Г.Н. Гистамин в биохимии и физиологии. — М.: Наука, 1981. — 274 с.
22 Переверзев В.А., Кубарко А.И., Балаклиевский А.И. и соавт. Роль серотонина и гистамина в повышении устойчивости организма к экстремальным воздействиям // Физиол. журн. им.Сеченова. — 1992. — Т.78. — № 6.
23 Джангозина Д.М. Гигиеническое обоснование метаболической концепции формирования и прогрессирования вибрационной болезни у шахтеров-угольщиков: Автореф. дис. ... д-ра мед. наук. — Караганда, 2003. — 48с.
24 Mohri K., Reiman H.L. Histamine content and mast cells in human gastric and duodenal mucosa // Agent and action. — 1978. — № 8. — P. 372-375.
25 Каценович Л.А., Ходжиев А.К., Джангозина Д.М. и соавт. О корреляции содержания гистамина и гистидина в крови, кининовой системы и состояния иммунологического гомеостаза у больных вибрационной болезнью. Актуальные вопросы гигиены и профессиональной патологии в условиях научно-технического производства. — Ташкент, 1980. — С. 241-243.
26 Смирнов В.С., Петленко С.В., Евстигнеев В.И. и соавт. К проблеме адаптации иммунной системы человека при экологической катастрофе // Военно-мед.журн. — 1992. — № 12. — С. 10-14.
27 Новицкий А.А., Успенская О.Н., Лесничий В.В. О механизме возникновения вторичных иммуннодефицитов в процессе хронического экологопрофессионального перенапряжения. Взаимодействие нервной и иммунной системы. — Оренбург, 1990. — С. 53, 54.