This research has been conducted to reveal the outcomes of rotation of crops in mountain and steppe districts near Almaty. The main morphogenetic characteristics have been investigated for the collection of representatives of common bean, Phaseolus vulgaris L. by using samples from various countries such as Kazakhstan, USA, China, Czech Republic, Poland, Turkey and Russia under different climate and soil conditions. A series of useful genetic stocks for the main economically valuable traits were detected. Several introduced varieties of common bean have showed high seed germination, maturation and significant resistance to water deficit, whereas some Kazakhstani varieties have been exceeded certain foreign specimens and varieties by weight of seeds and other parameters. It has been proven that “Luna” belonging to Czech collection would ripe the first (80 days from planting to complete technical maturity). Other varieties could approach the same conditions 10-12 days later. Using local specimen “Aktatti”, the effect of new bio organic mineral fertilizer produced by the Faculty of Chemistry has been demonstrated on morphogenetic traits of common beans. Basic catalogue of common bean resources including approximately 40 parental varieties and specimens of Phaseolus vulgaris L. and their relatives originally from different geographic regions has been compiled. It is planned to complete it by six bushy and runner representatives of French collection. The data have been completed by quantitative and qualitative amino acid evaluations. Kazakhstani and foreign cultivars and specimens respectively have been groupped by the data on amino acid composition of seeds which was analized by the method of liquid chromatography. Essential amino acids have approached approximately 30% of total amino acids pool in Kazakhstani specimens. If tyrosinylation index (Phe/Tyr ratio) for local specimens achieved 0.90 0.95, similar index for external varieties was equal to 0.88-0.89. This difference may accentuate the role and the quality of membrane proteins in local lines strengthening a composite stability (resilience, temperature and metabolic) in comparison with international common bean specimens under present study.
Introduction. A third of all food production is lost via leaky supply chains or spoilage. Food loss is a key contributor to global food insecurity. Resource-intensive animal-based food also limits food availability.
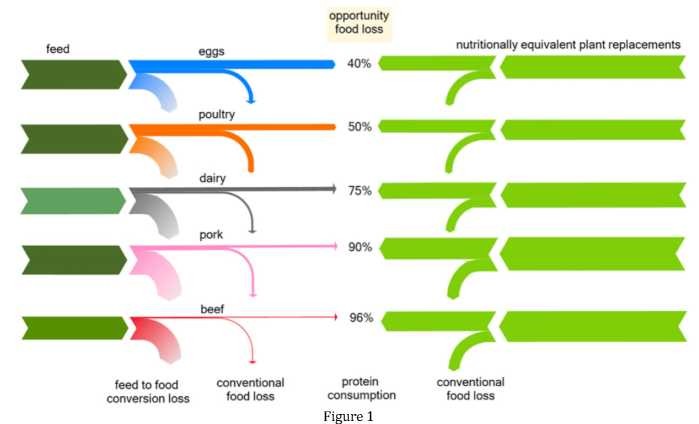
Shepon et al., 2018[1] have shown that plant-based replacements for basic animal categories in the US (beef, pork, dairy, poultry, and eggs) would produce from 2 to 20 times more nutritionally similar food per unit cropland. Replacing all animal-based items with plant-based replacement diets can add enough food to feed 350 million additional people, more than the expected benefits of eliminating all supply chain food loss.
The protein opportunity food loss is discussed by [1], from production to final consumption for the five major animal categories and their plant-based replacement diets. Each row shows the chain of protein from field to fork for each major animal category and its plant-based replacement diet. for the same land area used for all starting arrows.
Opportunity food loss values at consumption are given as percentages in the middle, demonstrating different protein contents between plant replacement diets (right) and the respective animal category (left) under identical land resource investments. For instance, the arbitrary land area needed to produce the feed for 4 g of beef protein will yield 100 g of protein of human-destined nutritionally equivalent plant diet, which is an opportunity food loss at the consumer level of 96% [1].
The process of cultivation of Phaseolus vulgaris L. populations or hybrids may be predicted from natural hybridization and natural selection of these forms which are adapted to changing climate conditions within the habitats. As a consequence crops would spread from these areas to new territories. Transformation of wild species keeps on extensively because of human activities, as the most valuable plants for feeding and life have been selected for the following cultivation [2]. During the process of evolution only plants with unique traits such as early germination and maturation, bush, type of growth, large or medium leaves and flowers, non-dehiscence and the absence of pergament layer in pods have been selected [3]. In comparison to other legumes, Phaseolus vulgaris L. is referred as more capricious to soil conditions due to its preference of light, fine grained clay-containing plastic soils or coarse grained, fertilized sand soils.
Cold and humid fine grained soils containing clay are not suitable to breed common bean. In addition, under high level of acidification, soil compaction and turf formation can also contribute to the negative effect of crops degradation [4]. Some heavy metals may significantly decrease production of common beans [5]. Due to acidic soils, the growth of nitrogen-fixing bacteria is suppressed. Usually it causes decreased amount of nitrogen in soil [6]. To improve yields of common bean, it is necessary to minimize acidic soils or add some calcium to them. Interestingly, there are a lot of varieties of common beans cultivated in Kosovo, in combination with Zea mays or as a monoculture [7]. It was determined that the stress had low impact on photosynthetic rates in common bean [8]. However, according to some authors, stress can influence on the process of photosynthesis in common beans.
Common bean, Phaseolus vulgaris L., should be mandatory included in the human diet due to high content of protein, vitamins, macroand microelements. А, C, E, РР, В, and К vitamins are natural anti-oxidants, strengthening the immune system, coordinating the body organs as promoting metabolic exchange processes. By protein content common bean is comparable with meat as it contains 21 g protein per 100g of the product. Arginine as abundant amino acid in the common bean composition, is in charge of nitrogen-exchanging processes thereby enabling significant reduction of sugar level in diabetis patients. Mineral salts based on iron, magnesium, phosphorus, iodine, potassium, sulphur and calcium are known to enhance body's struggle with bronchitis and gastrointestinal diseases. Common bean reveals diuretic effect and positive action on the urogenital system. Aminoacids lisine, tyrosine, and methionine were registered to facilitate quick protein acquisition thus alleviating bowel's activity. Common bean contains 300 kcal per 100 g of the seeds or frozen pods but is nevertheless regarded as essentially dietary product. It is regularly used in fasting diets. Owing to high protein concentration common bean is popular among vegetarians.
It is widely known that fresh common bean products conclude a range of toxins. Therefore, it is never recommended in kind of the raw, untreated courses. Toxins are deleted by the cooking, and then they are known to lose their danger. However, improper preparation of the common bean salad or dressing may cause the heaviest poisoning.
To summerize the list of the most studied medical effects caused by the common bean it is noteworthy to mention that it: is useful as antidiabetic food;
improves the activity of the alimentary tract; positively regulates cardiovascular system; sanifies the skin;
relieves the fatigue;
mitigates symptoms of rheumatoid arthritis; treats a cold;
prevents from Alzheimer's disease strengthens the bones;
displays anti-aging effect; treats cataract.
Contraindications to the use of the common bean concern patients facing high acidity, gastritis, cholecystitis, pancreatitis, ulcer, colitis, and gout.
Regardless of numerous types, common bean is one of the most useful food products. Red, white, black and green common beans are considered to possess distinct peculiarities and hence individual contraindications. However, green fresh and frozen, cooked or tinned beans do preserve the majority of beneficial features towards obligatory implication as a healthy food. (see open online resources: https://vsadu.ru/post/fasol-polza-vredsostav-kalorijnost-vidy.html; https: //chem-polezno.com/krupy/fasol and others).
Influence of the legume lectins is predominantly studied on metabolic and symbiotic traits of rhizobacteria (strains 634б, 646, 21110, 604к of Bradyrhizobium japonicum). Such symbiosis was determined under preliminary rhizobia incubation with purified lectins leading to increased peroxidase activity independent of potential symbiotic activity of the individual strain under investigation [9]. It was shown that lectins would cause the accumulation of new bacterial proteins to trigger nodule formation and nitrogen fixing capacity. Thus homolous lectins are able to induce multi-directional effect on genetically determined symbiotic potential of bacterial microsymbiont. Lectins were recently purified from the common bean seeds by conventional ammonium sulfate precipitation with their subsequent dialysis and molecular characterization using SDSPAGE [10]. Isolated lectins were shown to make up 31-34 kDa simultaneously demonstrating substantial hemagglutination activity across different human blood groups (A, B, AB and O). The agar-well diffusion technique has revealed the antimicrobial effect while measuring minimum inhibitory concentration (MIC). The data have indicated that lectins of all seeds under the trial would manifest a potent antibacterial activity to all the strains investigated (Staphylococcus aureus ATCC 6538, and Streptococcus mutants ATCC 25175, Pseudomonas aeruginosa ATCC 10145 and Klebsiella pneumonia), except that one (Escherichia coli 0157: HZ ATCC 51659) that remained unaffected. All lections under examination have disposed antifungal activity to Candida albicans. It was the first round of experiments implying pictures of scanning electron microscopy (SEM) to show the effect of saturated lectin fractions from common bean seed on Staphylococcus aureus and Pseudomonas aeruginosa. So the list of the medical common bean features mentioned above may be completed by evident antimicrobial and antifungal properties of extracted lectins.
Under specific conditions in mountain and steppe zone in the region of Almaty, morphogenetic traits of 37 representatives of Kazakhstani, American, Chinese, Czech, Polish, Russian, and Turkish collections have been assessed.
This experiment was conducted during two harvesting seasons in steppes and mountains. Both sites are located near the city of Almaty. 37 varieties and specimens were planted along with food and decorative pumpkins for the following purposes: i, to create and study Kazakhstani forms of common bean; ii, to create the collection to be explored by the university researchers and the students; iii, to develop the field and seed research capacities of Agrobiocenter at al-Farabi Kazakh National University; iv, to test composition of amino acids in different bean samples.
Material and methods. Current research was carried out during two seasons of field crop rotation in different regions such as mountain and steppe (plain) zones in the city of Almaty. 37 specimens of common bean and their relations were planted to accomplish the tasks described in the Introduction in details.
Part of stock specimens and varieties after preliminary propagation and introduction has been registered as the State Certificate on the subject of author rights No. 612 of 14 May, 2012 entitled “Distribution and exchange of bean specimens”. Varieties and specimens from the collection have been investigated according to protocols accepted by Awassa Agricultural Research Center and the Institute named after Vavilov[11]. Seeds twice were put in special plots of 10 x 2 meters, through the use of the method of row sowing with the distance approximately 40-60 cm between rows. “Aktatti”specimen was a standard one in specific regional conditions of Almaty. Our computer software entitled “Manager of panting” was used to design the experiment and plant all common beans.
Seeds of 17 cultivars and specimens of Phaseolus vulgaris L. were sown in the mountains. 20 cultivars and lines of common bean and its relatives (broad bean, Vicia faba L. and Turkish beans, Phaseolus coccineus L.) were planted in the steppe zone. Current work is concentrated on percentage of germination and length of pods and the content of amino acids in seeds. For this purpose the method of liquid chromatography was used. To increase germination we introduced 25-30g of domestic bioorganic fertilized per plot 25-35 days after planting with moderate watering.
Six representatives of domestic and foreign collections have been were selected to clarify composition of amino acids by highperformance liquid chromatography. Quantitative and qualitative analysis of amino acids was performed on the basis of acidic hydrolysis of extraction from seed powder in the presence of 6N HCl at 105oC for 24 hours. The hydrolysate was then dried 3 times with rotory evaporator at 40oC. The pellet obtained was resuspended with portions of sulfosalicylic acid and centrifuged to remove aggregated precipitates. Supernatant was exposed to liquid chromatography on the Dowex-50 coulmn at the elution rate equal to 150 l/sec. After neutralization of the column with deionized water, amino acid factions were washed out with 6N NH4OH (elution rate 300 l/sec).
The elate was dried with rotary evaporator under pressure of 1 atmosphere and 50-60°С. Mixture of SnCl2, 2,2-dimethyl oxypropan and propanol saturated with HCl was added to dried preparation. Then it was incubated at high temperature (to 110°С) for 20 min to be dried repeatedly. To conduct acetylation, dried pellet was treated with the mixture of acetic anhydride, triethanolamine and acetone (1:2:5). The mixture was incubated at 60°C for 1.5-2 min and dried. The preparation was diluted with ethyl acetate saturated with NaCl solution. The upper phase containing ethyl acetate, after thorough agitation, was picked out. Gas chromatography of amino acids in the decanted phase was carried out by using “Carlo Erba 4200”, Italy-USA. Amino acid elution was performed with a stainless-steel column (40 х 0.3 cm) filled with polar mixture of 0.31% Carbowax 20m, 0.28% Silar-5av and 0.06% Lexan on the Chromosorb WA-W-120-140m matrix. The standard regime of elution was maintained at the temperature of the flame ionisation detector 300°С, evaporator temperature of 250°С, initial column (furnace) temperature of 110°С and subsequent modes of column incubation: 6° per min in the range of 110-185° С and 32° per min in the range of 185250°. Maximum temperature of the column was supported until necessary fractions completely exited [12]. Statistic treatment of the data obtained was performed by the method of analysis of variance [13,14].
Results . Common bean seeds have been planted in the mountain zone in two sites independently. The first site is located at the territory of the Institute of Botany and Phytointroduction of the Science Committee of the Ministry of Education and Science of the Republic of Kazakhstan; the second site lies in Almarasan Gorge along the River Bolshaya Almatinka (800-1200 m above the sea level). The steppe plots are presented at the territory of “Zhanga Talap” centre proprietary of al-Farabi Kazakh National University and Kazakh Institute of Soil and Crop Research (both nearly 600 m above the sea level).
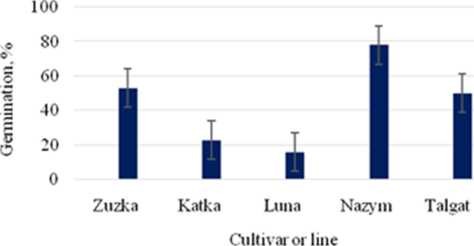
varieties and specimens of Phaseolus vulgaris L., have been sown on plots of the Institute of Botany and Phytointroduction; 8 specimens have been planted in the Almarasan Gorge (mountain zone). 20 varieties and specimens of common bean and its relatives (broad bean, Vicia faba L. and Turkish beans, Phaseolus coccineus L.) have been planted in the “Zhanga Talap” Agrobiocenter (steppe zone). Studies of morphogenic traits in the process of germination, showed that local Kazakhstani varieties and specimens surpass samples from Czech collection (see Figure 2). Among all Czech samples of common bean, cv. “Zuzka” at the 30th day after planting demonstrated the highest level of germination. Two other varieties “Katka” and “Luna”showed lower germination (23.3% and 16.6%, respectively). Also “Zuzka” has also been detected to interior other varieties and specimens by leaf size (11. 2 x 8.0 cm), whereas these parameters for “Katka” and “Luna” were 6.5 x 4.5 and 9.3 x 6.4 cm, respectively. At the same time “Zuzka” has approached the stage of flower formation earlier than other plants. The emergence value for “Zuzka” was much greater than percentages for other Czech common beans used for the experiment. This data has theoretical and evolutional significance because seed number and pods size, germination speed and lower length of stem in warm-seasonal legumes are construed as traits of domestication [15]. It has been revealed that common bean leaves possess oval or wide oval form under local conditions. In addition, some varieties and specimens revealed silver-polished stipules and different colour options for leaves, which is a genetic marker defining the variety. The colour is dependent on the vegetative stage of the plant, soil quality and amounts of fertilizers.
 Introduction and adaptation of foreign varieties to Kazakhstani climate conditions are in progress. One of the factors influencing on the positive dynamics is genetic polymorphism and possibility to adapt foreign samples, especially the representatives from Czech collection. In the mountains not far from the city of Almaty (site in the Almarasan Gorge) it has been established that three varieties such as “Zuzka”, “Katka”, and “Luna” (Figure 3) which were tried to be introduced in 2012, demonstrated high productivity with the exception of “Jitka”, which did not germinate at all.
Introduction and adaptation of foreign varieties to Kazakhstani climate conditions are in progress. One of the factors influencing on the positive dynamics is genetic polymorphism and possibility to adapt foreign samples, especially the representatives from Czech collection. In the mountains not far from the city of Almaty (site in the Almarasan Gorge) it has been established that three varieties such as “Zuzka”, “Katka”, and “Luna” (Figure 3) which were tried to be introduced in 2012, demonstrated high productivity with the exception of “Jitka”, which did not germinate at all.
Figure 2 - Results of phenological observations: intermediate evaluation of germination of representatives of Kazakhstani and Czech collections of common bean (30-th day after sowing)
a




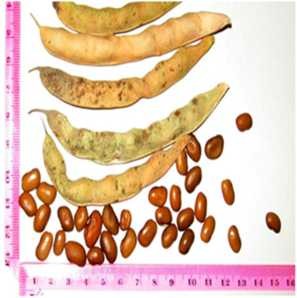

Seed material obtained from “Zuzka”, “Katka”, and “Luna” would be used for further breeding, at the territory of “Zhanga Talap” in the steppe zone. All Czech varieties have shown high resistance to pathogenic bean weevil (Acanthoscelides obtectus Sav.) at
room temperature. “Luna” became the earliest riping variety with a maturation period of 80 days from the onset of ontogenesis. Other specimens have approached the stage of technical maturity 10-12 days later.
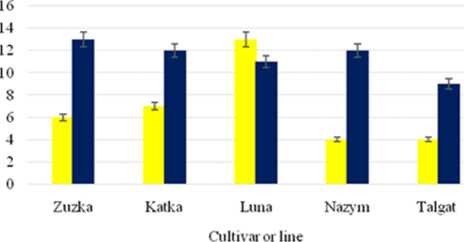
As we can see in the Fig.2, the maximal size of mature pod was obtained only on the 92-th day after planting, and belongs to “Zuzka” (13.3 + 0.1 cm). The pod size of “Katka” and “Luna” was approximately 12.0 + 0.2 cm and 10.8 + 0.1 cm, respectively. The domestic specimen “Aktatti” had similar pod length in the range of 11.0 + 0.1 cm, whereas other Kazakhstani representatives such as “Nazym” and “Talgat” had pod sizes of 12.4 + 0.1 and 9.0 + 0.2 cm, respectively during the final stage of technical maturity.
Pod length values of “Zuzka” and “Talgat” have reliably excelled the same parameter of other cultivars and lines under investigation (Figure 4).
Number of seeds per pod ■ Pod length. cm
Figure 4 Results of structural analysis of Kazakhstani and Czech varieties and specimens of common bean
Discussion. As seen from present study, preferentially suitable cultivars for the steppe zone are “Red Goya”, “Camelia”, “Bijchanka”, and “Ufimskaya”. While implication domestic “Aktatti” line positive impact of modern mineralized bioorganic fertilizer has been revealed on morphogenetic traits including productivity of Ph. vulgaris plants.
Current research is aimed at identification of superior lines obtained from a mountain location by phenotypic examination and comparing with exclusive Czech P. vulgaris cultivars. By the range of phenotypical characters (branch deviation, height, hypocotyl width, ripeness, lodging, disposition of upper pod, pod number per plant, and productivity[15] clusterization of available cultivars was carried out. In the Figure 5 shown percentage of emergence, which was processed using computational cluster analysis, domestic form “Nazym” is noticed to be neighbouring to Czech cv. “Zuzka”, whereas local line “Talgat” is being close to “Zuzka”, too. However, “Talgat” is recorded to be quite distant from two other Czech accessions, namely cvs “Katka” and “Luna”.
Figure 5 Comparison of common bean cultivars and lines clustered by the germination rate exhibit differences of Kazakhstan and
Czech collections of common bean (30-th day of sprouting) 1. Nazym, 2. Talgat, 3. Luna, 4. Katka, and 5. Zuzka.
Judging by the studies fulfilled, “Nazym” specimen may be attributed to a unique line due to morphogenetic characters. Data of Fig. 4 include local and Czech varieties and specimens, as the latters were successively introduced into the mountain zone. Noteworthy, Wang et al. proposed similar approach demonstrating by their studies genetic resemblance of the European and the North African genotypes which were closely associated with their geographical origins and ecological state (Wang et al., 2012). Another adequate research towards the enrichment and analysis of national bean collection was fulfilled by [16, 17]. Chinese investigators [19] attempted evaluating a collection of local samples for the genetic variability, genepool identity and relationships within and between the groups identified among the genotypes. The landraces in our reserch were clustered into two genepools. Earlier Polish researchers evaluated the genetic variability across commercial cultivars and domestic accessions of the dwarf and runner P. vulgaris types to reveal notable polymorphism among P. vulgaris and P. coccineus samples allowing to distinguish articulate sets of beans [20] One of advantageous outcomes of present investigation is the amount of polymorphism in stocks and “open gate” towards inducing foreign varieties, and the Czech collection in this particular case. Foothill experiments conducted in the Almarasan allowed to pick out three varieties such as “Zuzka”, “Katka” and “Luna” possessing a highly desirable traits such as high yield and early maturity for further research. It was noticed that Czech genotypes have exhibited high responsiveness to Acanthoscelides obtectus Sav. (bean weevil) under normal conditions. One of Czech representatives (“Luna”) was defined to be the earliest to reach maturity on 80-th day after planting. On the contrary, “Zuzka”and “Katka” were found to reach the same stage of maturity 10-12 days later. Amino acid composition has indicated a range of essential amino acids in domestic bean lines comprising approximately 30% of total amino acid quantities. Due to most significance of lysine and thryptophan for plant growth, it has been demonstrated that the the tyrosinylation index (TI), or Phe/Tyr ration across local lines would be fixed around 0.90-0.95, whereas TI for external cultivars would make up 0.88-0.89. This difference may accentuate the role and the quality of membrane proteins in local lines strengthening a composite stability (resilience, temperature and metabolic) incomparison with international common bean specimens under present study.
By the quantities of few amino acids (glutamate, aspartate, alanine, and proline) local bean samples have been revealed to outstrip external cultivars more than 2 times. The unique features of local bean specimen “Nazym” are confirmed by computational estimation of the amount of essential amino acids as seen from Figure 6.
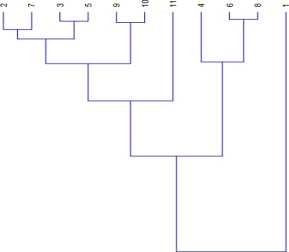
Figure 6 Comparison of common bean varieties and specimens clustered by the differences in essential amino acid contents in reference of Kazakhstani and
other international representatives 1. Nazym, 2. Talgat, 3. Luna, 4. Katka, 5. Zuzka, 6. Aktatti, 7. Bijchanka, 8. Camelia, 9. Dzhungarskaya,
10. Red Goya, and 11. Ufimskaya
We investigated the influence of newly elaborated bio organic mineral fertilizer on morphogenetic traits of local “Aktatti” bean accession. The data have manifested that the yield under a new fertilizer can be increased by 19-25%, regardless of climatic conditions.
Noteworthy, ongoing general progress in biology and biotechnology of legumes may be explained by tremendous diversification of the tree sub-branches in this research area. To specify this issue the authors have attempted to classify major trends in present legume studies as indicated in Table 1.
Analysis of morphogenetic traits across the available samples has led to compilation of the catalogue for main original varieties of P. vulgaris. The list includes approximately 40 parental specimens and their relatives of different initial origin. Out of the Czech bean varieties examined in the mountain zone, cv. “Luna” was detected to reach maturity the earliest (80 days of harvesting), whereas resting varieties have been recorded at technical maturity 10-12 days later. By germination rates verified by automated cluster test, the genotype “Nazym” was confirmed to be close by maturation period to cv. “Zuzka” and local speciment “Talgat”. “Nazym” has appeared to be quite promising for commercial production in the South-East of Kazakhstan. “Nazym” has also been ascribed to be advanced by essential amino acids, taking over the reference genotypes under current study.
Besides Czech and local P. vulgaris genotypes forementioned, six French commercial accessions of bushy and runner common beans have indicated high or average productivity in our conditions. Since cv. “Argus” is specific by its hybridization ability, it is obvious that the studies on bean domestication, crossing and molecular biology are essential. Further development of domestic bean collection along with the application of another eight directions of biology and biotechnology of legumes requires efforts both from researchers and authorities involved.
Noteworthy, ongoing general progress in biology and biotechnology of legumes may be explained by tremendous diversification of the tree sub-branches in this research area. To specify this issue the authors have attempted to classify major trends in present legume studies as indicated in Table 1.
|
N |
Research direction |
Citation(s) |
|
1 |
Bean domestication history and studies on orphan (underutilized) legumes |
Berrocal-Ibarra et al. 2002[21]; Cullis and Kunert. 2016[22]; Kwak et al. 2012[23]; Lopez Herrera et al. 2001[24]; Nanni et al. 2011[25]; Pena-Valdivia et al. 2010[26]. |
|
2 |
Bean plant physiology and biochemistry; food legume productivity research; combined studies on dryland cereals and legumes |
Daryanto et al. 2015[27]; Hyman et al. 2016[28]. |
|
3 |
Bean genetics and chromosome biology |
Varao Vasconcelos et al. 2015[29]. |
|
4 |
Bean molecular biology; Bean RNA biology |
Hiz MC et al. 2014[30] Kavas et al. 2014[31]. |
|
6 |
Bean virology; genomoviral studies |
Silva Lamas et al. 2016 [32]. |
|
7 |
Bean symbiotic studies and bean pathology |
Ballhorn et al. 2016 [33]. Marcenaro D and Valkonen JPT, 2016 [34]. |
|
8 |
Bean metabolic engineering and biofortification |
Ramirez Rivera et al. 2016 [35]. |
|
9 |
Bean dietology; bean nutrigenetics |
Gomez Ojeda et al. 2015 [36]. |
|
1 |
Bean volatiles research; bean signalomics |
Zhang et al. 2012 [37]. |
Table 1 List of research trends in common bean bi
and bi

Analysis of morphogenetic traits across different accessions has allowed to compile the basic catalogue of original P. vulgaris cultivars and lines. It includes more than 40 parental specimens and their relatives of different origin.
Cv. “Luna” was detected to reach maturity the earliest (2.5 months), out of the Czech bean varieties examined in the mountain zone. Remaining genotypes have achieved maturation a fortnight later.
According to cluster distribution by germination rates, local line “Nazym” has been proven to be close by maturation period to cv. “Zuzka” and local line “Talgat”. Local line “Nazym” has appeared to be quite promising to be grown commercially in South-East regions of Kazakhstan for ornamental and nutritional aims. This line is also referred to be advanced by the essential amino acids content, surpassing the reference cultivars and lines under present study.
Six French commercial accessions of bush and runner common beans have revealed high or average productivity in our conditions. Since cv. “Argus” is specific by its hybridization ability, it is obvious that the studies on bean domestication, crossing and molecular biology are essential. Further development of domestic bean collection for the application of another eight directions of bean biology and biotechnology requires efforts both from researchers and authorities involved.
REFERENCES
- Shepon A., Eshelb G., Noorc E., Milo R. The opportunity cost of animal based diets exceeds all food losses // Proc Natl Acad Sci. 2018. №115(15). Р. 3804-3809.
- Bodnar G.V., Lavrinenko G.T. Grain legume crops. M.: Kolos, 1977. 256 p.
- Zhukovsky PN. Crops and their relations. Leningrad: Kolos, 1971. 791 р.
- Popov V.P, Martynov O.L. Morphological and biological peculiarities of few cultivars for common bean in the South of Moscow Region // Proceedings of Russian Acad. Agric. Sci. 2001. №4. Р. 21-23.
- Ratushnyak A.Y, Ratushnyak A.A., Andreeva M.G, Kayumov A.R., Bogachev M.I., Trushin MV Effect of lead and salicylic acid on some plant growth parameters in Pisum sativum L. // World Applied Sciences Journal. 2012. №19(8). Р. 1157-1159.
- Zerfus V.M, Schitov A.G, Kozlova GYa Factors determining the formation of symbiotic apparatus and its impact on grain legumes productivity in Western Siberia // Agrochemistry. 1997. №12. Р. 27-31.
- Fetahu S, Aliu S, Rusinovci I, Kelmendi B, Maliqi N Accumulation and distribution of dry matter in some common bean landrace seedlings at the phase of the cotyledons // Agriculture & Forestry. 2013. №59. Р. 133-142.
- Shekari F., Javanmard A., Hassanpouraghdam M. Response of two red bean (Phaseolus vulgaris L.) cultivars to controlled water deficit stress during post —flowering growth stage // Agriculture & Forestry. 2014. №60(1). Р. 245-257.
- Sytnikov DM, Kruglova E.D., Mandrovskaya N.M. Soybean lectin effect on metabolism and symbiotic properties of Bradyrhizobium japonicum strains // Biotechnology. 2011. №4. Р. 642-650.
- Hamed EE, Ibrahim MME, Mervat M, Antimicrobial Activities of Lectins Extracted from Some Cultivars of Phaseolus vulgaris Seeds // J. Microb Biochem Technol. 2017. №9(3). Р. 109-116.
- Korsakov N.I. et al. Methodical Instructions on The Study of The Collection for Grain Legume Crops. All Union Institute of Plant Research. Leningrad: 1975. 59 р.
- Adams R. Determination of amino acid profiles in biological samples by gas chromatography // J. Chromatography. 1988. №431(2). Р. 271-284.
- Dospekhov B.A. Field experiment techniques (with fundamentals of statistic treatment of research data). M.: Agropromizdat, 1985. 351 р.
- Bisgaard S (2008) Must a Process be in Statistical Control before Conducting Designed Experiments? // Quality Engineering, ASQ. 2008.№2. Р. 143-176.
- Isemura T., Kaga A., Konishi S., Ando T., Tomooka N., Han O.K., Vaughan D.A. Genome dissection of traits related to domestication in azuki bean (Vigna angularis) and comparison with other warm —season legumes // Ann. Bot. 2007. №100(5). Р. 1053 -1071.
- Beattie AD, Michaels TE, Pauls KP Predicting progeny performance in common bean (Phaseolus vulgaris L.) using molecular markerbased cluster analysis // Genome. 2003. №2. Р. 259 -267.
- Asfaw A, Blair MW, Almekinders C Genetic diversity and population structure of common bean (Phaseolus vulgaris L.) landraces from the East African highlands // Theor. Appl. Genet. 2009. №1. Р. 1-12.
- Kumar V., Sharma S., Sharma A.K., Kumar M., Sharma S., Malik S., Singh K.P., Sanger R.S., Bhat K.V. Genetic diversity in Indian common bean (Phaseolus vulgaris L.) using random amplified polymorphic DNA markers // Physiol. Mol. Biol. Plants. 2008. №14(4). Р. 383387.
- Zhang X, Blair MW, Wang S. Genetic diversity of Chinese common bean (Phaseolus vulgaris L.) landraces assessed with simple sequence repeat markers // Theor. Appl. Genet. v. 2008. №117(4). Р. 629-640.
- Nowosielski J, Podyma W, Nowosielska D Molecular research on the genetic diversity of Polish varieties and landraces of Phaseolus coccineus L. and Phaseolus vulgaris L. using the RAPD and AFLP methods // Cell Mol. Biol. Lett. 2002. №7(2B). Р. 753-762.
- Berrocal -Ibarra S., Ortiz -Cereceres J., Peca -Valdivia C.B. Yield components, harvest index and leaf area efficiency of a sample of a wild population and a domesticated variant of the common bean Phaseolus vulgaris // South African Journal of Botany. 2002. №68. Р. 205-211.
- Cullis C. and Kunert K.J. Unlocking the potential of orphan legumes // Journal of Experimental Botany. 2016. №68(8). Р. 1-9.
- Kwak M, Toro O, Debouck DG, Gepts P Multiple origins of the determinate growth habit in domesticated common bean (Phaseolus vulgaris) // Annals of Botany. 2012. №110(8). Р. 1573-1580.
- Lopez Herrera M, Aguirre Rivera J.R., Trejo C., Pena —Valdivia C.B. Differences in seed germination of wild and domesticated common bean (Phaseolus vulgaris L.) in response to storage // South African Journal of Botany. 2001. №67. Р. 620-628.
- Nanni L., Bitocchi E., Bellucci E., Rossi M, Rau D, Attene G, Gepts P., Papa R. Nucleotide diversity of a genomic sequence similar to SHATTERPROOF (PvSHP1) in domesticated and wild common bean (Phaseolus vulgaris L.) // Theor. Appl. Genet. 2011. №123. Р. 1341-1357.
- Pena -Valdivia C.B., Sanchez -Urdaneta A.B., Meza Rangel J., Juarez Munoz J., Garda -Nava I.R., Celis R. Velazquez Anatomical root variations in response to water deficit: wild and domesticated common bean (Phaseolus vulgaris L.) // Biol. Res. 2010. №43. Р. 417427.
- Daryanto S., Wang L., Jacinthe P. A. Global Synthesis of Drought Effects on Food Legume Production // PLOS ONE. 2015. №10. Р. 1371— 1374.
- Hyman G., Barona E., Biradar C., Guevara E., Dixon J., Beebe S., Castano S.E., Alabi T., Gumma M.K., Sivasankar S., Rivera O., Espinosa H., Cardona J. Priority regions for research on dryland cereals and legumes // Version 2.F1000Res. 2016. №5. Р. 885-889.
- Varao Vasconcelos E., Fellipe de Andrade Fonseca A., Pedrosa —Harand A., Cilene de Andrade Bortoleti K., Benko -Iseppon A.M., Felix da Costa A., Brasileiro -Vidal A.C. Intra and interchromosomal rearrangements between cowpea [Vigna unguiculata (L.) Walp.] and common bean (Phaseolus vulgaris L.) revealed by BAC -FISH // Chromosome Res. 2015. №23. Р. 253-266.
- Hiz M.C, Canher B., Niron H., Turet M. Transcriptome Analysis of Salt Tolerant Common Bean (Phaseolus vulgaris L.) under Saline Conditions // PLOS ONE. 2014. №9(3). Р. 259-267.
- Kavas M., Baloglu M.C., Atabay E.S., Ziplar U.T., Dasёgan H.Y., Uenver T. Genome-wide characterization and expression analysis of common bean bHLH transcription factors in response to excess salt concentration // Mol. Genet. Genomics. 2016. №291. Р. 129-143.
- Silva Lamas N., Salgado Fontenele R., Lucas Melo F., Felix Costa A., Varsani A., Graca Ribeiro S. Complete Genome Sequence of a Genomovirus Associated with Common Bean Plant Leaves in Brazil // Genome Announcements. 2016. №4(6). Р. 124-133.
- Ballhorn D.J., Schadler M., Elias J.D., Millar J.A., Kautz S. Friend or Foe —Light Availability Determines the Relationship between Mycorrhizal Fungi, Rhizobia and Lima Bean (Phaseolus lunatus L.) // journal.pone. 2016. №10. Р. 269-275.
- Marcenaro D, Valkonen JPT Seedborne Pathogenic Fungi in Common Bean (Phaseolus vulgaris cv. INTA Rojo) in Nicaragua // PLoS ONE. 2016. №11(12). Р. 147-153.
- Ramirez Rivera N.G., Garcia —Salinas C., Aragao F.J.L., Diaz de la Garza R.I. Metabolic engineering of folate and its precursors in Mexican common bean (Phaseolus vulgaris L.) // Plant Biotechnology Journal. 2016. №14. Р. 2021-2032.
- Gomez O., Wrobel K, Corrales Escobosa AR, Torres Elguera JC, Garay —Sevilla Ma E., Molybdenum and Copper in Four Varieties of Common Bean (Phaseolus vulgaris) New Data of Potential Utility in Designing Healthy Diet for Diabetic Patients // Biol. Trace Elem. Res. 2015. №163(1-2). Р. 244-254.
- Zhang S., Wei J., Kang L. Transcriptional Analysis of Arabidopsis thaliana Response to Lima Bean Volatiles // PLoS ONE. 2012. №7(4). Р. 226-234.