One of the earliest published examples of zooarchaeology was written by paleontologist Chester Stock in 1929 [1; 281–289]. He was given an assortment of animal bones found in the La Brea Tar Pits in Southern California. Instead of simply publishing a list of species, he identified the type of bone as known as an element, and calculated the minimum number of individual animals that must have fallen into the tar pit to have created this specific collection of bones. Stock’s analysis of the bones beyond simple identification of species is one of the earliest examples of modern techniques of zooarchaeology. Zooarchaeologists identify more than just species, and are able to analyze the number of animals or the amount of meat that was consumed on an archaeological site. Common data that is recorded off of a typical archaeological bone includes the species, element, side, sex, age, condition of the bone, type of breakage, and evidence of butchery, dog gnawing or artifact working. Contemporary zooarchaeology can now answer questions about how animals were exploited, for example, if they were raised for wool, milk, or meat, if they were butchered using stone or metal tools, or if they were ritually deposited in specific contexts.
CONTEXT
The example discussed here is a very small collection of caprine bone, which was found in the summer of 2011 by I.A.Kukushkin. The archaeological site is a Bronze Age kurgan which is located in Karkaraly National Park. This kurgan contains multiple graves and artefacts which are associated with the Andronovo culture. The collection was found buried just above a human grave, and was bagged separately from the other animal bones found in the kurgan.
Initial assessment of the bone identified signs of weathering, or exposure to sun and wind, as evidenced by their fragile nature and the longitudinal cracking along the shafts. The collection was highly fragmented due to its fragile nature. Species identification to caprine (or a category that includes sheep and goat) was done using bone identification books and articles [2–6]. Mathematical measurements to determine the exact species were done using digital calipers and the method outlined in Boesneck 1969, which is discussed below [7; 331–358]. Additional information about tooth identification and wear also came from an assortment of books and articles [8–11] and information about ageing bones came from Silver 1969 [12; 283–302].
RESULTS AND DISCUSSION
Most of the bone in this collection is highly fragmented, and as a result, not identifiable to species. There were 77 bones which could be identified, and 82 teeth which could also be identified. The remaining 471 fragments were too small to be classified into species of type of bone. Of the identifiable bone, all the bones were from the feet of caprines. The bone surface had degraded from exposure to the weather, and unfortunately identification of cut marks was not possible. Furthermore, identification for marrow extraction from the feet bones, which can be identified by the pattern of breakage of the bones, was obscured by the weathering fractures which had broken the bones into multiple pieces. However, there was still quite a lot of data that could still be found from this collection.
Species and Age
In this report, the bones have been described as ‘caprine’ which is a term that combines both sheep and goat. The bones of sheep (Ovis aries) and goat (Capra hircus) are notoriously difficult to differentiate, and the differences between these two species vary not in definite ways which can be easily spotted, but instead in very subtle ways, which can change depending on the region of study and the breed of sheep or goat. The most accepted way to differentiate the two is to compare all of the caprine bones within an assemblage and to use modern reference examples and published illustrations [5,6,7].Additionally, there are quantitative methods which can also differentiate between sheep and goat. Boesneck 1969 describes a measurement method utilizing the metapodia using a simple pair of calipers [7]. By taking two measurements on the distal medal condyle of the metapodia and creating a ratio, it is easy to determine which bones are sheep (those which have ratio <.63 are sheep, those >.63 are goat).
In this collection, the ratio of the measurements on the metapodia were all well into the sheep range of measurement. Additionally, most of the bones matches with the illustrations of sheep bones in the literuature. Interestingly, the first phalanges of these sheep did exhibit a robust shape which tend to be characteristic of goats. However, the overwhelming presence of sheep characteristics on the other foot bones and the measurement ratio which was well into the range of sheep confirmed this as a collection of sheep feet.
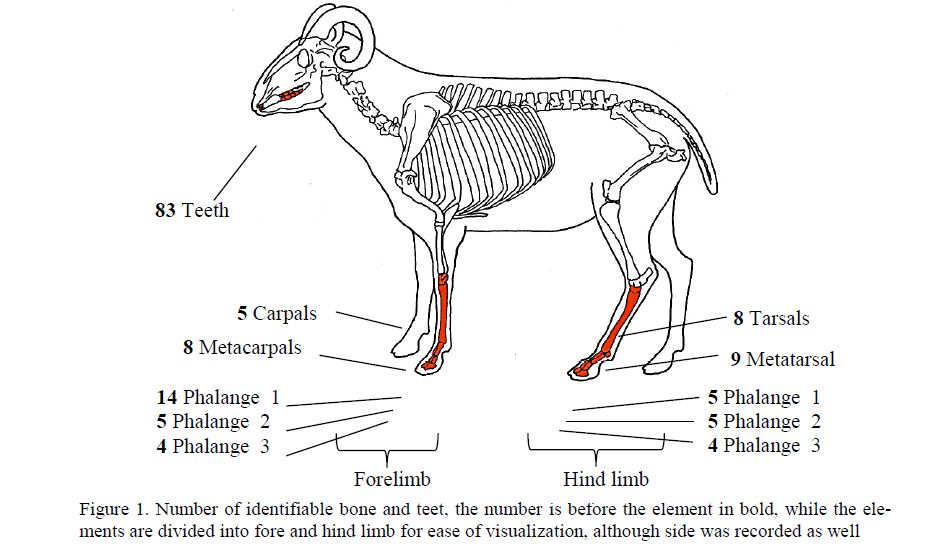
Figure 1. Number of identifiable bone and teet, the number is before the element in bold, while the elements are divided into fore and hind limb for ease of visualization, although side was recorded as well
To determine the age of the sheep, it is fairly simple to record the fusion data on the bones. Long bones in mammals grow towards their epiphysis (ends) and only fuse to these epiphysis at known times. If the bone is whole, and there are no unfused ends on the bones, the animal can be assumed to have reached the age of fusion before it was killed. This gives an age range after which the animal must have been slaughtered. In this collection, all of the metapodia were fused except for one example. Sheep metapodia fuse between 18– 28 months of age, so it can be assumed that these were adult sheep when they were slaughtered.
How Many Sheep? How Many Legs?
The first question asked: how many caprines were buried here seems like a simple one. However, counting the number of animals deposited is not as simple as it seems. The bones in an archaeological site do not represent entire animals, but instead reflect a collection of animal bones that were deposited. Unlike human bone, animals are rarely buried intact. Instead, animal bone is often deposited as a unit, for example as a leg, or even as a single bone, such as from a kitchen.
The simplest way to count bones is called NISP, or the Number of Identified Specimens. This is a simple count of the identifiable bone fragments and teeth which can be identified to species and element. NISP is a very basic unit of counting, however NISP can overestimate the presence of a species or an element for collections of bone that are very fragmented. In this example, the NISP is 159 (77 bones, and 82 teeth). In order to take into account fragmentation, another counting method, called MNE, or Minimum Number of Elements, is used. MNE is very similar to NISP, except that it only tallies bone fragments that can be identified to side, and takes into account bone fragments that may have originally formed a single bone [13; 214]. MNE is a simple way to factor out fragmentation and to look at the numbers of elements that were most likely deposited in the context. This counting method is shown below in Figure 2.
There are other counting methods to estimate the amount of meat or the number of individual animals that created a collection. The method called MAU, or Minimum Animal Units, was invented by Binford [14] in order to quantify how modern day Nunamuit people were selecting and transporting parts of reindeer from the kill site to their camp site. MAU works by adding together the left and right long bone fragments (except for the phalanges and other bones in the body which do not occur uniformly across species, which is discussed more thoroughly in Lyman 2008) and all of these fragments are added together to give an index numberor unit numberfor that species. This method is used for comparing different species within different collections within and across archaeological sites. MAU is useful for comparing which parts of animals were consumed and deposited in the site. This kind of analysis can reveal social divisions, as aristocrats most likely ate different parts of the animals, which were considered tastier, than the poorer settlers. In our example, MAU is not particularly useful, as we only have one species and one collection, however the calculations for MAU are given below in Figure 2.
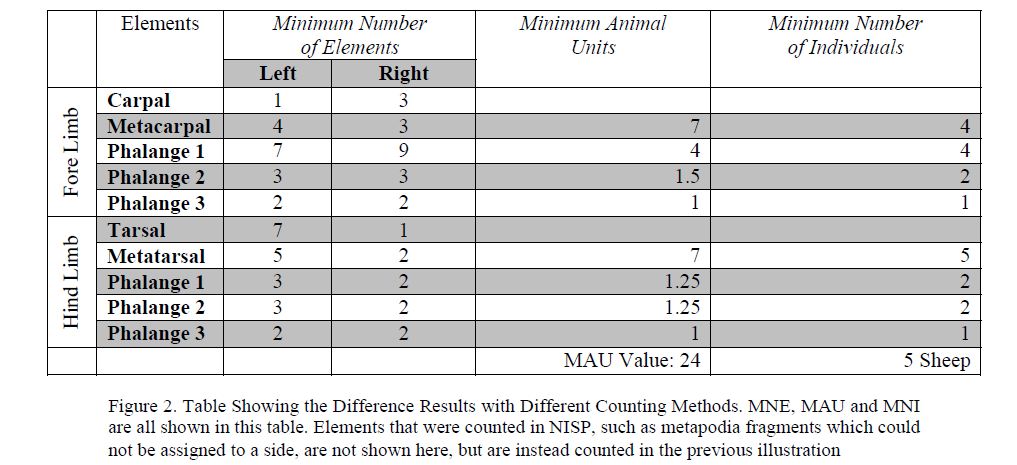
Figure 2. Table Showing the Difference Results with Different Counting Methods. MNE, MAU and MNI are all shown in this table. Elements that were counted in NISP, such as metapodia fragments which could not be assigned to a side, are not shown here, but are instead counted in the previous illustration
In special cases where there have been intact animal skeletons revealed, a counting method called MNI, or Minimum Number of Individuals, is very useful for calculating the number of animals that were slaughtered. This method was mentioned above, when Chester Stock invented it in his investigation of the tar pits at Rancho La Brea [1]. Stock took the MNE and simply used the most frequent side of each element, and then picked the most frequent element to calculate the minimum number of individuals. This theory is based upon the basic idea that there can be only one left long bone, for example, one left humerus, that each animal can contribute to a collection. If there are three left humeri, and two right humeri in specific context, then there must have been at least three animals that were killed. This method, while evocative, as it seems to easily give a good idea of the number of animals that were killed, only works in contexts in which the entire animal carcass was deposited. As animal bone is frequently butchered, and pieces are distributed and deposited in different contexts, it is not useful to use this calculate except for carcass burials. In Figure 2, the MNI is calculated to show the difference between the different calculations, however as we know that there were only feet and teeth found in this context, MNI is not the correct counting method to utilize.
As we can see from the example in Figure 2, the different counting methods all give very different results. While the MNI is very convincing in suggesting that 5 sheep were killed, it is important to remember that carcasses were not found in this depositinstead it was just feet, which are animal units. However, the MAU, while helpful in calculating a value in units, is not helpful here as there are no other deposits or species to compare with. Instead, the most useful calculation we have is the MNE. This simple calculation displays exactly what bones were found, while controlling for fragmentation. If you control for duplicate bones which occur more than once (such as the phalanges and the carpals and tarsals) it is clear that there were 4 left fore limbs, 3 right fore limbs, 5 left hind limbs, and 2 right hind limbs.
Teeth
These counting methods are not generally used on teeth, largely due to their number, the presence of deciduous dentition and the difficulty in siding and describing whether teeth are from the jaw or from the skull. In this case, teeth were roughly described to mandible or maxilla (skull), and it was found that there was a few teeth that were deciduous, but most were adult. This matches with the fusion data, which suggested that there may have been one juvenile slaughtered. Tooth wear patterns, which can be used to find the precise age of sheep, is only useful when there are entire sets of teeth, either in the jaw, or in the skull, so as to make an assessment of all of the teeth together [11]. Figure 3 shows a rough estimate of the number of jaws and skull sides that were deposited.

Figure 3. A rough count of the numbers of mandibles and maxillas present in the assemblage, calculated on teeth that could be identified and sided
CONCLUSION
It is possible to conclude, having explored all of the different suggestions, that there were 4 left fore limbs, 3 right fore limbs, 5 left hind limbs, and 2 right hind limbs which were placed into this context. We also know that there were at least 3 skulls and 4 jaws from the teeth. These were sheep, which had rather unusual goat characteristics on their first phalanges but in all other cases had very strong sheep morphology. It is most likely that a number of sheep, perhaps even as many as 5, were slaughtered and all of the other bones, which have quite a lot of meat, were taken away to be consumed in a different place. Interestingly, there were no astragali found. Astragalus are part of the ankle joint, and while other bones from that joint were found in this context, there were no astragali. This is most likely due to the fact that they have been used in dice games for milenia, and these bones were probably cut out of the joint before the bones were deposited.
It is unclear whether this is a ritual deposit or an offering. The location on top of a grave is very suggestive of a ritual deposit, however without analogies from other kurgans, it is hard to be certain. As these bones do not have as much meat as other parts of the sheep, it is also possible that this is just a deposit of butchery refuseor parts of the animal that were left behind after slaughter.
References
- Stock A Census of the Pleistocene Mammals of Rancho La Brea, Based on the Collections of the Los Angeles Museum // Mammology. — № 10. — 1929. — P. 281–289.
- Barone R. Anatomie comparee des mammiferes domestiques. — Paris: Vigot Freers, 1976. — 428
- Gilbert B.M. Mammalian Osteo-Archaeology: North America. — Columbia: Missouri Archaeological Society, 1973. — 337
- Hillson Mammal Bones and Teeth: An Introductory Guide of Methods of Identification. — London: Institute of Archaeology, UCL, 1999. — 64 p.
- Prummel W. & Frisch H.-J. A Guide for the Distinction of Species, Sex and Body Side in Bones of Sheep and Goat // Archaeological Science. — № 13. — 1986. — P. 567–577.
- Zeder A. & Lapham H.A. Assessing the reliability of criteria used to identify postcranial bones in sheep, Ovis, and goats, Capra // Archaeological Science. — № 37(11). — 2010. — P. 2887–2905.
- Boessneck J. Osteological differences between sheep (Ovis aries Linné) and goat (Capra hircus Linné) / D.Brothwell & Higgs (Eds.) Science in Archaeology: A Survey of Progress and Research. London: Thames and Hudson, 1969. — P. 331–358.
- Halstead , Collins P. & Isaakidou V. Sorting the Sheep from the Goats: Morphological Distinction between the Mandibles and Mandibular Teeth of Adult Ovis and Capra // Archaeological Science. — № 29. — 2002. — P. 545–553.
- Payne Morphological distinctions between the mandibular teeth of young sheep, Ovis, and goats, Capra // Archaeological Science. — № 12(2). — 1985. — P. 139–147.
- Zeder A. & Pilaar S.E. Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats, Capra // Archaeological Science. — № 37. — 2010. — P. 225–242.
- Payne S. Kill-off patterns in sheep and goats: the mandibles from Asvan Kale // Anatolian Studies. — № 23. — 1973. — P. 281–303.
- Silver A. The ageing of domestic animals. In (D.Brothwell & E.Higgs, Eds) / Science in Archaeology. 2nd ed. — London: Thames and Hudson, 1969. — P. 283–302.
- Lyman R.L. Quantitative Paleozoology. — Cambridge: Cambridge UP, 2008. — 348
- Binford L.R. Nunamit ethnoarchaeology. — New York: Academic Press, 1978. — 509