В статье отмечено, что полиморфные варианты генов, связанных с детоксикацией и поддержанием редокс-баланса организма, могут быть ассоциированы с индукцией активных радикалов и оксидатив-ным стрессом, возникающим в ответ на облучение. Представлен анализ распределения частот генотипов и аллелей по полиморфным локусам генов CAT, SOD2, SOD3, NOS, CYP1A1, GSTP1 в группе лиц, подвергшихся хроническому радиационному воздействию. Также приведен анализ частоты встречаемости этих генотипов в группах по полу и этнической принадлежности.
Введение
Немаловажную роль в реализации последствий действия ионизирующего излучения на организм играют механизмы оксидативной защиты. Одним из эффектов воздействия ионизирующего излучения является возникновение свободных радикалов [1, 2], которые могут являться индукторами окси-дативного стресса. Индукция оксидативного стресса в клетке посредством взаимодействия многих путей и компонентов может изменять палитру экспрессии генов и определять дальнейшую судьбу клетки [3]. Избыток радикалов может приводить к одиночным и множественным оксидативным повреждениям ДНК, таким как повреждения отдельных оснований, одно- и двунитевые разрывы ДНК. Эти повреждения в случае неэффективной репарации могут привести к гибели клетки или стать причиной мутаций, генетической нестабильности, малигнизации клетки [4]. Кроме повреждения ДНК, радикалы способны ингибировать ферменты, окислять липиды мембран (запуская перекисное окисление липидов)[5], тем самым нарушая функциональную активность клеток [6].
Поддержание баланса между процессами образования и утилизации активных радикалов — ре-докс-баланса — важный аспект жизни клетки и организма в целом. Клетки поддерживают редокс-баланс при помощи различных агентов, утилизирующих свободные радикалы, таких как глутатион, витамины С и Е, ферменты-антиоксиданты каталаза, супероксиддисмутаза, пероксидаза [7]. Одно-нуклеотидные полиморфизмы в генах, кодирующих факторы, участвующие в поддержании редокс-баланса, могут влиять как на степень экспрессии генов, так и на функциональную способность соответствующих ферментов.
Фермент каталаза (CAT) выполняет функцию защиты клеточных компонентов от перекиси водорода, разлагая ее с образованием водорода и воды. Полиморфизм rs7943316 гена CAT выражается в замене -21A/T в промоторе гена, предположительно влияет на экспрессию данного гена.
Митохондриальная супероксиддисмутаза (SOD2) — фермент, дисмутирующий супероксид-анион в форму пероксида водорода, который впоследствии элиминируется ферментом CAT. Полиморфный вариант T201C (rs4880) гена SOD2 выражается в аминокислотной замене, которая нарушает альфа-спираль белка SOD2; мутантный белок имеет сниженную на 30-40 % активность, увеличивая подверженность клетки оксидативному стрессу [8].
SOD3 — внеклеточная супероксиддисмутаза, оказывает защитное действие относительно окислительного повреждения ДНК и канцерогенеза у млекопитающих [9]. Полиморфизм C896G(rs1799895) связан с повышенным уровнем SOD3 в плазме крови [10].
CYP1A1 кодирует один из белков семейства цитохромов P450. Он относится к монооксигеназам, метаболизирует ксенобиотики, такие как полициклические ароматические углеводороды, содержащиеся в табачном дыме. Образующиеся промежуточные окисленные метаболиты способны реагировать с белками и нуклеиновыми кислотами [11], что может стать началом злокачественного процесса в клетке. Полиморфизм rs1048943 гена CYP1A1 приводит к аминокислотной замене Ile462Val в каталитическом центре фермента [12] и к продукции энзима с повышенной активностью [13]. Показано, что присутствие в геноме высокоактивного аллеля гена CYP1A1 приводит к значимому увеличению частоты клеток с хромосомными аберрациями [14].
Большое число исследований говорит об ассоциации возникновения онкологических заболеваний с нарушениями функционирования глутатион S-трансфераз — ферментов-антиоксидантов, использующих глутатион в реакциях трансформации широкого спектра метаболитов и продуктов окси-дативного стресса [15, 16]. Аминокислотная замена Ile105Val гена GSTP1 выражается в пониженной активности фермента [17].
Оксид азота (NO) — свободный радикал, выступающий в роли медиатора в таких процессах, как нейротрансмиссия, регуляция давления крови, антимикробная и противоопухолевая активность [1]. В организме синтезируется из L-аргинина ферментами NO-синтазами: в нейрональных клетках — NOS1, в клетках-макрофагах — NOS2, в эндотелиальных клетках — NOS3 (eNOS). При взаимодействии с радикалами кислорода в условиях оксидативного стресса NO может быть источником образования пероксинитрита. Известно, что минорный аллель гена NOS3 по полиморфизму rs1799983, выраженному в аминокислотной замене Asp298Glu, ассоциирован с различными расстройствами сердечно-сосудистой системы [18-20].
Таким образом, учитывая важную роль генов детоксикации ксенобиотиков и генов, вовлеченных в ответ на оксидативный стресс, в развитии радиационно-индуцированных эффектов актуальным является изучение распределения частот их аллелей по полиморфным локусам.
Материалы и методы
Общая обследуемая группа состояла из 172 человек, куда вошли люди, подвергшиеся хроническому радиационному воздействию в 1950-1960 гг. в результате загрязнения радиоактивными отходами ПО «Маяк» бассейна реки Течи [21]. Население прибрежных сел подверглось комбинированному внешнему (преимущественно у-) и внутреннему у- и Р-радиационному воздействию с низкой ЛПЭ. Основными источниками внешнего у-облучения являлись: вода, донные отложения и пойменные почвы, а также участки территории населенных пунктов, загрязненные в результате хозяйственной деятельности (поливные огороды, механический перенос радиоактивности). Внутреннее облучение было обусловлено радионуклидами, поступавшими в организм с речной водой и продуктами питания местного производства (молоко, рыба, овощи с поливных огородов). Наиболее высокие уровни суммарной Р-активности были обнаружены в рыбе и мясе водоплавающей птицы. Менее загрязненными смесью радионуклидов были молоко и овощи [22].
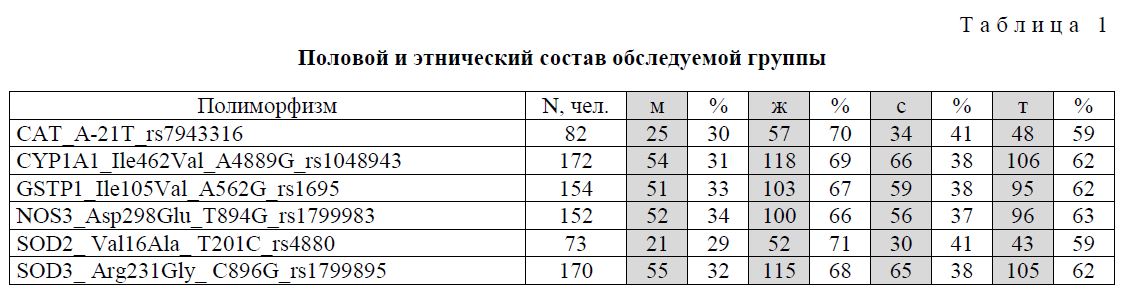
Основным дозообразующим радионуклидом являлся 90Sr, который, будучи аналогом кальция, накапливался и долгое время удерживался в костной ткани. Кроме того, значимый вклад в формирование дозы внесли 137Cs и 89Sr, но, благодаря короткому периоду полураспада 89Sr и быстрому выведению из организма 137Cs, облучение за счет этих радионуклидов реализовывалось только в первые 5 лет после их поступления [23]. Все обследованные имели индивидуальную реконструированную дозу на красный костный мозг, средняя доза для общей группы составила 1,08±0,06 Гр (от 0,0005 до 4,69 Гр). Характеристика группы по половому и этническому составу представлена в таблице 1.
Геномная ДНК была выделена из замороженной цельной венозной крови, хранящейся в банке тканей ФГБУН УНПЦРМ. Часть образцов была выделена с помощью набора реагентов iPrep Pure Linkg DNAB lood Kit на приборе Applied Biosystems Library Builder, а часть — методом фенол-хлороформной экстракции согласно описанной ниже методике. После размораживания 700 мкл крови переносили в пробирку эппендорф и добавляли 700 мкл ASSC. Центрифугировали при 12000 об./мин 2 минуты. К осадку добавляли 1400 мкл ASSC, перемешивали и вновь центрифугировали при 12000 об./мин 2 минуты. К осадку добавляли 270 мкл ацетатного буфера, 30 мкл 10 % SDS, инкубировали при 37 °С 1 час. Затем проводилась очистка фенол-хлороформной смесью (50/50). После центрифугирования (12000 об./мин 10 минут) надосадочную жидкость переносили в чистую пробирку и осаждали ДНК 1000 мкл спирта (100 %). После кратковременного центрифугирования проводили отмывку 70 %-ным спиртом. Получившийся осадок ДНК высушивали и растворяли в 100 мкл деио-низированной воды.
Генотипирование проводилось методом аллель-специфичной ПЦР «в реальном времени» с использованием наборов фирмы «Ген-Эксперт» (Москва, РФ) на амплификаторе «AppliedBiosystems StepOne™ Plus» (LifeTechnologies™, США). Анализ данных генотипирования проводился с использованием программ StepOne™ Softwarev2.1, TaqMan® GenotyperSoftwarev1.3. Для сравнения полученных в эксперименте частот аллелей с частотами европейской и азиатской популяций использовались данные интернет-ресурсов HapMap (http://hapmap.ncbi.nlm.nih.gov/), 1000 GenomesProject (http://www.1000genomes.org/), dbSNP (http://www.ncbi.nlm.nih.gov/snp). Проверка соответствия распределения генотипов закону Харди-Вайнберга осуществлялась путём сравнения ожидаемых и наблюдаемых частот генотипов с использованием критерия ^2.Анализ ассоциации генотипов с этнической принадлежностью (ЭП) и полом проводился с использованием коэффициента %2 по Пирсону, статистический анализ — с помощью программ Microsoft Excel (Microsoft Corporation, 2010), STATISTICA v10 (StatSoft, Inc., 2011).
Результаты и обсуждение
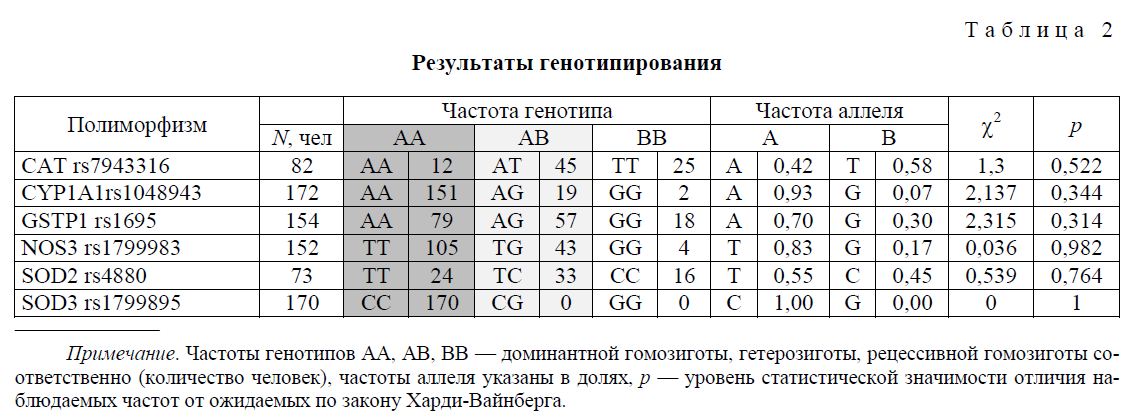
Результаты генотипирования образцов ДНК исследуемой группы людей представлены в таблице 2. Проверка соответствия распределения аллельных вариантов закону Харди-Вайнберга не выявила статистически значимых отклонений, что может свидетельствовать об отсутствии факторов, способствующих отбору по данным полиморфным аллелям.
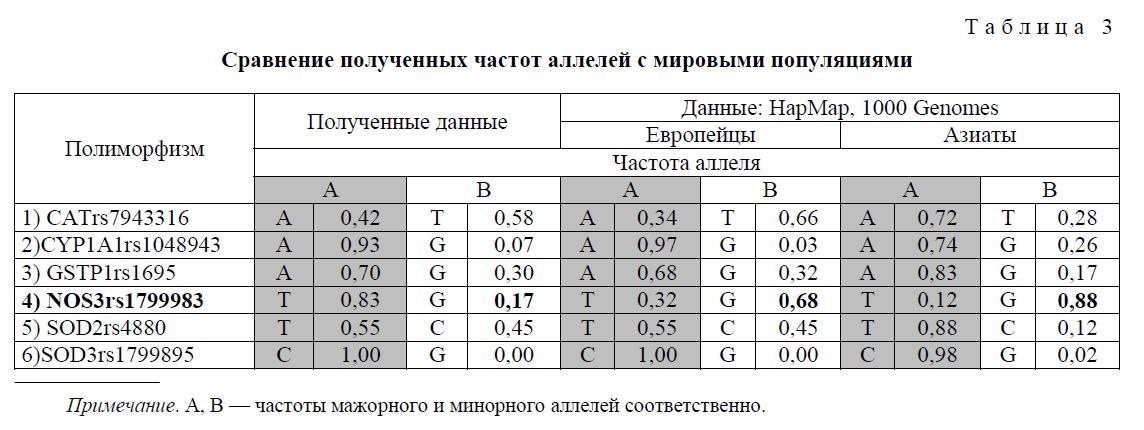
В таблице 3 представлены результаты сопоставления полученных аллельных частот с частотами в европейской и азиатской популяции.
Из данных таблицы 3 видно, что в большинстве случаев частоты аллелей в обследованной группе соответствуют «европейским» частотам. Для полиморфизма NOS3 частота минорного аллеля значительно понижена (0,17 в исследованной группе против 0,68 и 0,88 в европейской и азиатской популяциях соответственно).
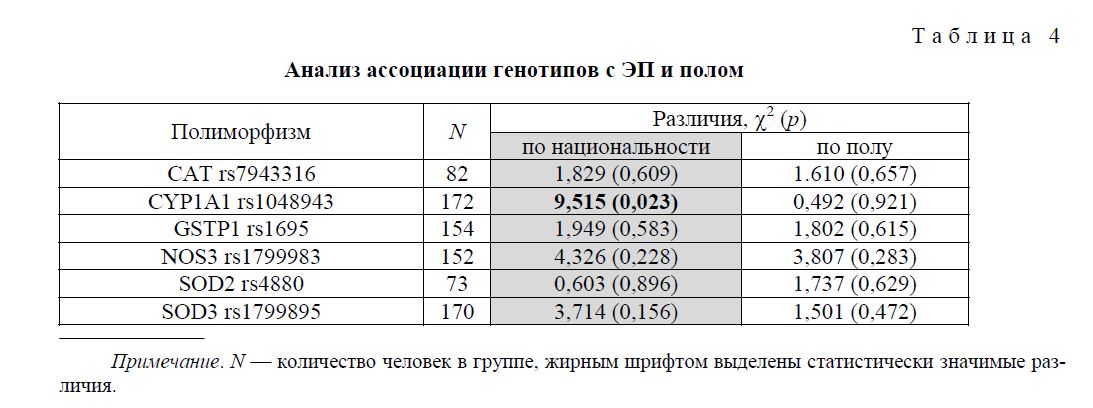
Также нами был проведен анализ ассоциации генотипов по изучаемым полиморфным локусам с полом и национальностью обследованных людей, который показал достоверное различие в распределении генотипов по локусу CYP1A1 rs1048943 между славянами и тюркитами: для славян частоты гомозигот по мажорному аллелю, гетерозигот и гомозигот по минорному аллелю были соответственно: 95,4 %, 4,6 %, 0 %, а для тюркитов: 83 %, 15,1 %, 1,9 %. Данные результаты согласуются с результатами исследования [11], в котором также наблюдалось увеличение носительства минорного аллеля среди тюркитов (башкир).
Заключение
В результате проведенного исследования нами были получены частотные распределения для генотипов и аллелей по полиморфным локусам CAT A-21T rs7943316, CYP1A1 Ile462ValA4889 Grs1048943, GSTP1 Ile105ValA562Grs1695, NOS3 Asp298GluT894Grs1799983, SOD2 T201CVal16 Alars4880, SOD3 C896G Arg231Gly rs1799895 в изучаемой группе. Не было выявлено отклонений наблюдаемых частот генотипов от равновесного распределения Харди-Вайнберга, что свидетельствует об отсутствии факторов отбора. Была выявлена ассоциация аллеля CYP1A1462Val с принадлежностью к этнической группе тюркитов.
Список литературы
- Владимиров Ю.А. Свободные радикалы в биологических системах // Соросовский образовательный журнал. — 2000. — Т. 6, № 12. — С. 13-19.
- Стюарт Ф.А. и др. Отчет МКРЗ по тканевым реакциям, ранним и отдаленным эффектам в нормальных тканях и органах — пороговые дозы для тканевых реакций в контексте радиационной защиты / Ред. А.В.Аклеев, М.Ф.Киселев; Перев. Е.М.Жидкова и Н.С.Котова. — Челябинск: Книга, 2012. — С. 248-249. (Тр. МКРЗ; публ. 118). — 384 с.
- Klaunig J.E., Kamendulis L.M., Hocevar B.A. Oxidative stress and oxidative damage in carcinogenesis // Toxicol. Pathol. — 2010. — 38, No. 1. — P. 96-109.
- Kryston T.B., Georgiev A.B., Pissis P., Georgakilas A.G. Role of oxidative stress and DNA damage in human carcinogenesis // Mutat. Res. — 2011. — 711, No. 1-2. — P. 193-201.
- Avery S. V. Molecular targets of oxidative stress // Biochem. J. — 2011. — 434, No. 2. — P. 201-210.
- Limdn-Pacheco J., Gonsebatt M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress // Mutat. Res. — 2009. — 674, No. 1-2. — P. 137-147.
- Скулачев В.П. Кислород в живой клетке: добро и зло // Соросовский образовательный журнал. — 1996. — № 3. —С. 4-10.
- Sutton A., Imbert A., Igoudjil A., Descatoire V., Cazanave S., Pessayre D., Degoul F. The manganese superoxide dismutase Ala16Val dimorphism modulates both mitochondrial import and mRNA stability // Pharmacogenet. Genomics. — 2005. — Vol. 15, No. — P. 311-319.
- Singh B., Bhat H.K. Superoxide dismutase 3 is induced by antioxidants, inhibits oxidative DNA damage and is associated with inhibition of estrogen-induced breast cancer // Carcinogenesis. — 2012. — Vol. 33, No. 12. — P. 2601-2610.
- Sandstrom J., Nilsson P., Karlsson K., Marklund S.L. 10-fold increase in human plasma extracellular superoxide dismutase content caused by a mutation in heparin-binding domain // J. Biol. Chem. — 1994. — Vol. 269. — P. 19163-19166.
- Кочетова О.В., Корытина Г.Ф., Ахмадишина Л.З. и др. Анализ полиморфизма гена цитохрома Р450 1А1 (CYP1A1) в этнических группах Республики Башкортостан // Генетика. — 2008. — Т. 44, № 12. — С. 1677-1683.
- Hayashi S.-I., Watanabe J., Nakachi K., Kawajiri K. PCR detection of an A/G polymorphism within exon 7 of the CYPlAl gene // Nucleic Acids Res. — 1991. — 19, No. 17. — P. 4797.
- Crofts F., Taioli E., Trachman J. et al. Functional significance of different human CYP1A1 genotypes // Carcinogenesis. — 1994. — 15, No. 12. — P. 2961-2963.
- Дружинин В.Г., Волков А.Н., Головина Т.А., Глушков А.Н., Минина В.И., Ларионов А.В., Мейер А.В., Лунина А.А., То-лочко Т.А. Роль молекулярного полиморфизма генов репарации и биотрансформации в оценке радиочувствительности человека к воздействию излучений радона // Актуальные проблемы токсикологии и радиобиологии: Тез. докл. Рос. науч. конф. с междунар. участием. СПб., 19-20 мая 2011 г. — СПб.: Фолиант, 2011. — 312 с.
- Fryer A.A., Bianco A., Hepple M., Jones P.W., Strange R.C., Spiteri M.A. Polymorphism at the glutathione S-transferase GSTP1 locus. A new marker for bronchial hyperresponsiveness and asthma // Am. J. Respir. Crit. Care Med. — 2000. — Vol. 161, No. — Р. 1437-1442.
- Valko M., Leibfritz D., Moncol J., Mark T.D. Cronin, Milan Mazur, Joshua Telser. Free radicals and antioxidants in normal physiological functions and human disease // The International Journal of Biochemistry & Cell Biology. — 2007. — 39. —44-84.
- Aguiar E.S., Giacomazzi J., Schmidt A.V., Bock H., Saraiva-Pereira M.L., Schuler-Faccini L., Duarte Filho D., dos Santos P.A., Giugliani R., Caleffi M., Camey S.A., Ashton-Prolla P. GSTM1, GSTT1, and GSTP1 polymorphisms, breast cancer risk factors and mammographic density in women submitted to breast cancer screening // Rev. Bras. Epidemiol. — 2012. — Vol. 15,No. 2. — P. 246-255.
- Berger K., Stogbauer F., StollM., Wellmann J., Huge A., Cheng S., Kessler C., John U., Assmann G., Ringelstein E.B., Funke The glu298asp polymorphism in the nitric oxide synthase 3 gene is associated with the risk of ischemic stroke in two large independent case-control studies // Hum. Genet. — 2007. — Vol. 121. — P. 169-178.
- Casas J.P., Bautista L.E., Humphries S.E., Hingorani A.D. Endothelial nitric oxide synthase genotype and ischemic heart disease: meta-analysis of 26 studies involving 23028 subjects // Circulation. — 2004. — 109. — P. 1359-1365.
- Jachymova M., Horky K., Bultas J., Kozich V., Jindra A., Peleska J., Martasek P. Association of the glu298-to-asp polymorphism in the endothelial nitric oxide synthase gene with essential hypertension resistant to conventional therapy // Biochem. Biophys. Res. Commun. — 2001. — 284. — P. 426-430.
- Аклеев А.В. Медико-биологические и экологические последствия радиоактивного загрязнения реки Теча / Аклеев А.В., Киселев М.Ф. — М.: Медбиоэкстрем, 2000. — 532 с.
- Дегтева М.О., Воробьева М.И., Толстых Е.И., Шагина Н.Б., Кожеуров В.П. Дозиметрическая система реки Теча: реконструкция доз для оценки риска радиационных последствий // Вопросы радиационной безопасности. — 2000. — № 4. — С. 36-46.
- Tolstykh E.I., Degteva M.O., Peremyslova L.M., Shagina N.B., Shishkina E.A., Krivoshchapov V.A., Anspaugh L.R., Napier B.A. Reconstruction of long-lived radionuclide intakes for Techa riverside residents: strontium-90 // Health Physics. — 2011. — 101. — P. 28-47.