В статье с использованием методов микробной экологии (индексы сходства Жаккара и Раупа-Крика, анализ главных координат) выявлены ассоциации микроорганизмов в ожоговых ранах. Показано, что устойчивые ассоциации могут сформироваться в результате антибиотикотерапии и антагонистических взаимоотношений между экологическими группами микроорганизмов. Представлены экспериментальные данные, дано их обсуждение.
Воздействие высокой температуры при ожогах пламенем и при контакте с термическим агентом способно «стерилизовать» рану. По мере пребывания больного в стационаре ожоговая рана контами-нируется микрофлорой, при этом скорость заселения зависит от многих факторов: места получения травмы, способа оказания первой помощи, локализации ожога, наличия госпитальных возбудителей и т.д. [1].
По результатам исследований, проведённых нами ранее, приоритетными микроорганизмами, выделяющимися из ран пациентов Челябинского областного ожогового центра, являлись S. aureus, P. aeruginosa и A. baumannii [2]. Высокая этиологическая значимость данных микроорганизмов отмечается и в работах других исследователей, что позволяет считать их основными возбудителями ожоговой инфекции [3-5]. При этом основные возбудители часто встречаются в ассоциациях с различными условно-патогенными микроорганизмами: E. coli, Proteus sp., Klebsiella sp. и др. [6-8]. Закономерности формирования подобных ассоциаций остаются недостаточно изученными. Вместе с тем выявление причин экологической общности разных микроорганизмов в ожоговых ранах важно для каждого стационара, так как позволяет определить стратегию антибиотикотерапии и противоэпи-демиологического режима. Поэтому цель данной работы заключалась в выявлении устойчивых ассоциаций микроорганизмов в ожоговых ранах и объяснении возможных причин их экологической общности.
Материалы и методы
Исследование проводилось на базе бактериологической лаборатории МБУЗ ГКБ № 6 г. Челябинска. Были проанализированы результаты бактериологических посевов пациентов, госпитализированных в Челябинский областной ожоговый центр с января по июнь 2012 г. Всего исследована 341 проба раневого отделяемого.
Для выявления ассоциаций микроорганизмов в ходе статистического анализа использовали расчёт индексов сходства Жаккара Cj и Раупа-Крика CR-C, а для интерпретации ассоциаций полученные матрицы индексов сходства подвергали многомерной ординации методом главных координат. Оценку статистической значимости для Cj проводили с использованием таблиц критических значений [9], а для CR-C рассчитывали как P = 1 - CR-C.
Статистически значимыми признавали ассоциации микроорганизмов при P < 0,05, незначимыми — при P > 0,10; в промежуточных случаях (0,05 < P < 0,10) обсуждали тенденции к ассоциациям. Расчёты и графические построения выполнены в пакете PAST (v. 2.17c) [10].
Результаты и обсуждение
Из 341 пробы в 104 (30,5 %) посев раневого отделяемого не дал роста, т.е. высеваемость микрофлоры составила 69,5 % (n = 237). В 176 пробах (51,6 %) возбудитель был представлен монокультурой. В 61 пробе (17,9 %) были выделены ассоциации из 2 и более микроорганизмов, из них 29 проб (48,3 %) содержали смешанные популяции приоритетных микроорганизмов: S. aureus, P. aeruginosa и A. baumannii.
В результате анализа 61 пробы с ассоциациями были рассчитаны показатели экологической общности, которые представлены в таблице.
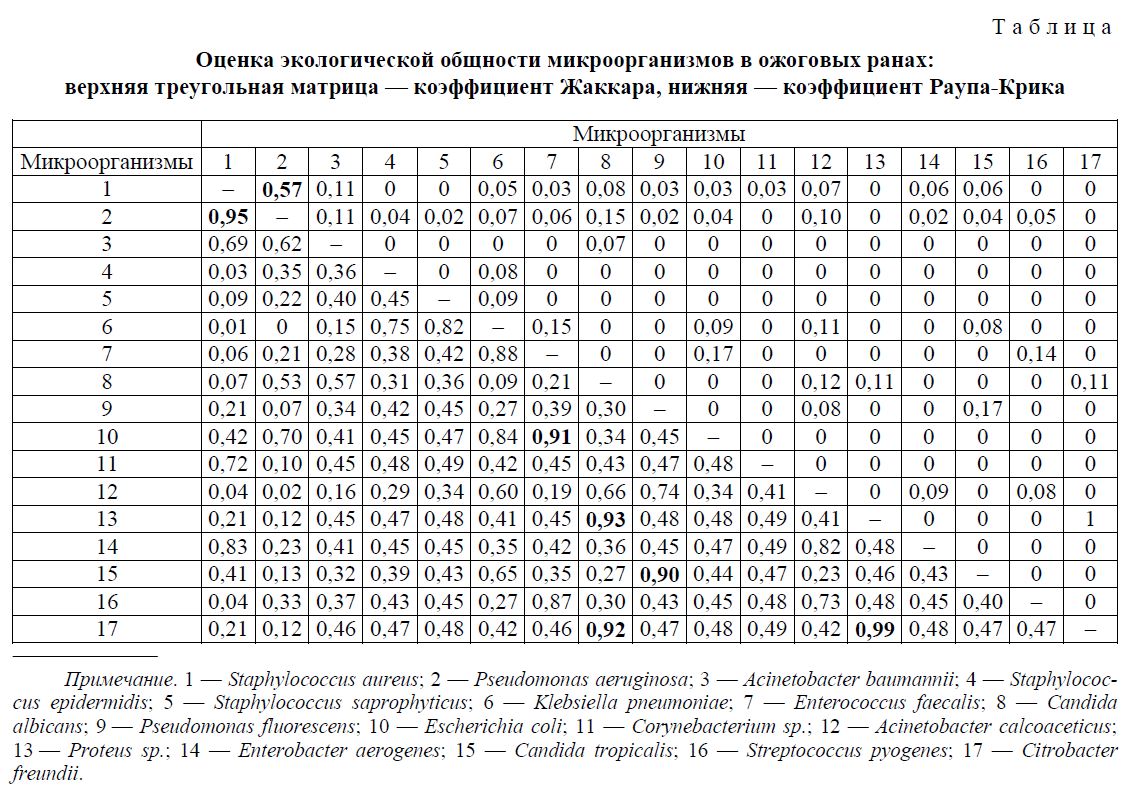
Коэффициент Жаккара Cj, наряду с коэффициентом Дайса (Сёренсена, Чекановского-Сёренсе-на), является одной из наиболее распространённых парных мер сходства в фаунистических, флористических, экологических исследованиях и применяется также в микробной экологии [11]. В контексте данной работы он представляет собой отношение числа проб, содержащих одновременно два микроорганизма, к числу проб, содержащих хотя бы один из этих двух микроорганизмов. Таким образом, Cj = 0 указывает на отсутствие совместной встречаемости организмов, а Cj = 1 — на исключительно совместную встречаемость двух организмов. С помощью данного коэффициента удалось выявить только одну статистически значимую ассоциацию: S. aureus и P. aeruginosa (Cj = 0,57; P = 0,001).
Коэффициент Раупа-Крика CR-C является вероятностной мерой сходства. Он рассчитывается с использованием рандомизационной процедуры Монте-Карло на основе сравнения наблюдаемого числа случаев совместного присутствия организмов в пробе с распределением их совместной встречаемости в 1000 случайным образом сгенерированных выборках из данных по обоим видам. Поскольку CR-C представляет собой вероятность неслучайной ассоциации организмов, оценку статистической значимости для него можно вычислить как P = 1-CR-C. С помощью данного коэффициента удалось выявить 2 статистически значимые ассоциации: Proteus sp. и Citrobacter freundii (CR-C = 0,99; P = 0,008) и S. aureus и P. aeruginosa (CR-C = 0,95; P = 0,048). Также была обнаружена тенденция к ассоциациям следующих микроорганизмов: C. albicans и Proteus sp. (CR-C = 0,93; P = 0,069), C. albicans и Citrobacter freundii (CR-C = 0,92; P = 0,080), Enterococcus faecalis и E. coli (CR-C = 0,91; P = 0,092), Pseudomonas fluorescens и Candida tropicalis (CR-C = 0,90; P = 0,098).
Для визуализации обнаруженных ассоциаций и объяснения возможных причин экологической общности микроорганизмов, относящихся к разным таксонам, использовали многомерный анализ.
Анализ главных координат (многомерное метрическое шкалирование) представляет собой одну из техник непрямого градиентного анализа в экологии [12], родственную анализу главных компонент. Если в качестве меры сходства между показателями используется корреляция Пирсона, то результаты анализа главных координат будут аналогичны методу главных компонент. Однако в анализе главных координат можно использовать и другие меры сходства (в нашем случае — индексы Жаккара, Раупа-Крика и др.), тогда как выделенные в его результате оси (главные координаты, ГК) интерпретируются аналогично главным компонентам.
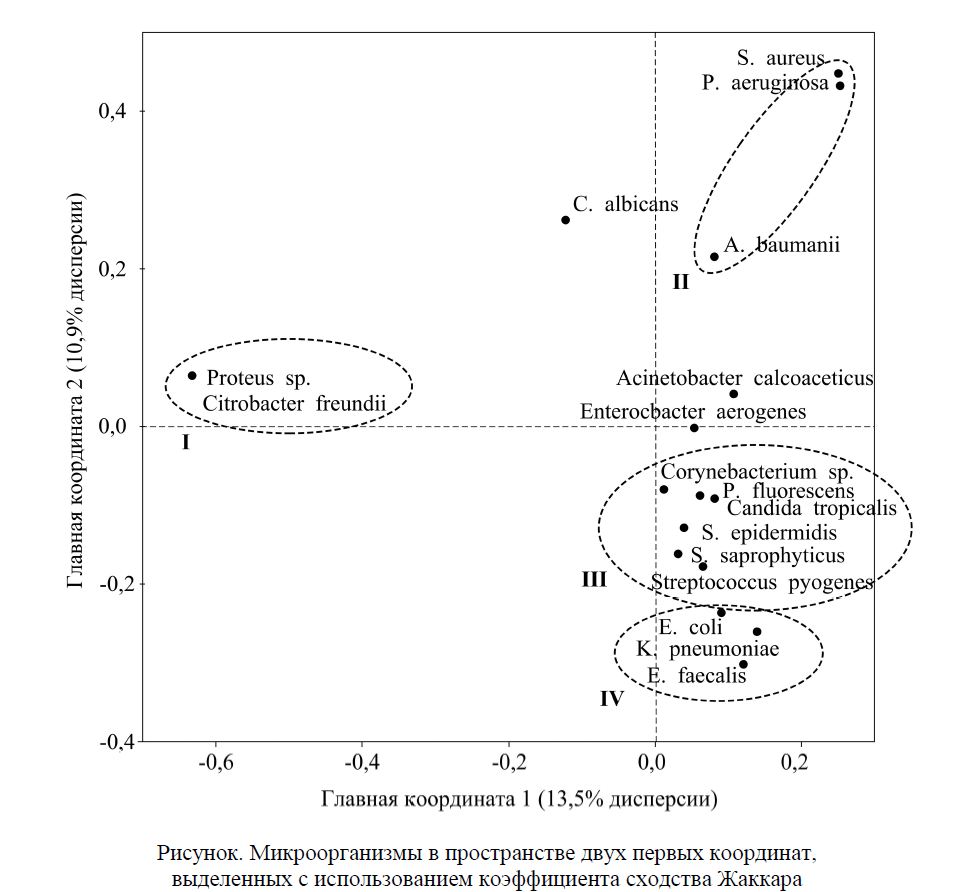
В ходе анализа главных координат было установлено, что данные слабо подвергались обобщению: даже на первую главную координату (ГК 1) приходилось только 13,5 % общей изменчивости совместного распределения микроорганизмов. Это указывает на существование большого числа факторов, специфически влияющих на микробный пейзаж.
На рисунке таксоны микроорганизмов представлены в пространстве двух первых координат, выделенных с использованием индекса сходства Жаккара (результаты с использованием индекса Раупа-Крика аналогичны и не приводятся). Объекты, находящиеся по разные стороны от нулевого значения, обнаруживают отрицательную связь, по одну сторону — положительную. Из рисунка видно, что все исследованные таксоны микроорганизмов распределились на 4 основных группы, для которых были характерны высокие коэффициенты экологической общности (обведены пунктиром). В группу I вошли представители кишечной микрофлоры, в группу II — приоритетные возбудители ожоговой инфекции, III — представители кожной микрофлоры, IV — предположительно, госпитальная флора фекального происхождения.
Вдоль ГК 1 проявилась отрицательная связь группы I со всеми остальными. Вошедшие в неё Proteus sp. и Citrobacter freundii являются представителями кишечной микрофлоры, которая заселяет поверхность ожоговых ран в первые дни после ожога [1]. Таким образом, ГК 1, вероятно, отражает временную динамику смены микробного пейзажа, когда первичные контаминанты раны замещаются на госпитальную микрофлору. Следовательно, выявленная устойчивая ассоциация Proteus sp. и Citrobacter freundii может маркировать пробы, собранные на ранних сроках госпитализации.
Вдоль ГК 2 отчётливо разделились группы, включающие представителей кожной микрофлоры (III), госпитальной микрофлоры фекального происхождения (IV) и приоритетных возбудителей ожоговой инфекции (II). Полагаем, что ГК 2 отражает смену микробного пейзажа в процессе анти-биотикотерапии. Действительно, стандартная схема ведения ожоговых больных предполагает использование Р-лактамных антибиотиков в качестве стартовой терапии. Эти антибиотики эффективно подавляют в первую очередь кожную микрофлору (группа III), но остаются неэффективными в отношении микроорганизмов группы IV [13]. Поэтому на следующем этапе лечения пациентам назначают карбапенемы, которые ингибируют рост представителей группы IV. В результате такого лечения в ожоговой ране создаётся оптимальная среда для размножения устойчивых к Р-лактамам и кар-бапенемам микроорганизмов, сформировавших группу II. Таким образом, логичное объяснение получает устойчивая ассоциация Enterococcus faecalis и E. шһ. Её представителей объединяет резистентность к Р-лактамам: для E. faecalis характерна природная устойчивость к антибиотикам, обусловленная наличием хромосомной кассеты генов полирезистентности, а для E. — приобретённая, обусловленная наличием Р-лактамаз расширенного спектра. Наиболее сильная и высоко статистически значимая ассоциация S. aureus и P. аeruginosa также формируется в результате антибиоти-котерапии, оставляющей в ране полирезистентные штаммы указанных бактерий. Это объясняет высокий процент высеваемости именно этих видов микроорганизмов на протяжении ряда лет [14]. Интересной особенностью является близость Candida albicans к экологической группе II. Этот факт можно объяснить тем, что при использовании системной антибактериальной терапии грибы занимают освободившуюся от бактерий экологическую нишу.
Не исключено, что взаимоотношения между выделенными группами микроорганизмов определяются не только антибиотикотерапией, но и являются результатом антагонизма. Так и у S. aureus, и у P. аeruginosa установлена способность продуцировать бактериоцины, ингибирующие рост представителей кишечной и кожной микрофлоры. В свою очередь для E. шһ, K. pneumoniae и C. freundii также характерна способность к бактериоциногении. Считается, что выход бактериоцинов зависит, прежде всего, от показателей плотности культуры-продуцента и далее регулируется собственной сигнальной системой — чувством кворума (quorum-sensing systems, Qs). Чувство кворума позволяет популяциям бактерий коллективно регулировать экспрессию генов и синхронизировать поведение группы, являясь своего рода «нейронной» сетью бактерий [15]. Таким образом, нерациональная ан-тибиотикотерапия может привести к избыточному размножению антибиотикорезистентных штаммов, которые не только способны вызвать инфекцию, но и за счет массовой секреции бактериоцинов подавить рост популяции других бактерий, занимающих эту же экологическую нишу.
Выводы
- С использованием методов микробной экологии в ожоговых ранах выявлены устойчивые ассоциации микроорганизмов, причем на долю популяции приоритетных микроорганизмов(S.aureus, P. aeruginosaиA. baumannii)приходилось 48,3 % от всех ассоциаций.
- Представители разных экологических общностей редко обнаруживались совместно, что может быть следствием антимикробной терапии и/или свидетельством антагонистического характера взаимоотношений между исследованными группами бактерий.
Список литературы
- Алексеев А.А., Крутиков М.Г., Яковлев В.П. Ожоговая инфекция. Этиология, патогенез, диагностика, профилактика и лечение. — М.: Вузовская книга, 2010. — 416 с.
- Андреева С.В., Бахарева Л.И., Нохрин Д.Ю. Видовой состав микрофлоры ожоговых ран // Вестн. Челябинского гос. ун-та. — 2013. — № 7 (298), Биология, Вып. 2. — С. 58-59.
- Keen E.F. Incidence and bacteriology of burn infections at a military burn center // Burns. — 2010. — 36. — P. 461468.
- Kiuru V., Anttila V., Vuola J. Eradication of Acinetobacter baumannii Outbreak at a Burn Centre // Supplement to Journal of Burn Care and Research. — 2012. — 33, No. 2. — P. 15.
- WongТ.Н., Tan B.H., Ling M.T., SongС.Multiresistant Acinetobacter baumanii on a burns unit — clinical risk factors and prognosis // Burns. — 2002. — 28, No. 4. — P. 340-349.
- Шамаева С.Х., Миронов С.Х., Потепов А.Ф., Петрова К.М., Евграфов С.Ю. Микрофлора ожоговых ран и её чувствительность к антибиотикам у детей с ожоговой болезнью // Человек и его здоровье: Курский науч.-практ. вестн. — 2011. — № 2. — С. 91-95.
- Iman A. Hussien, Khalid A. Habib, Kifah A. Jassim. Bacterial Colonization of Burn Wounds // J. Baghdad for Sci. — — No. 4. — P. 623-631.
- Nagoba B.S., Deshmukh S.R., Wadher B.J., Pathan A.B. Bacteriological analysis of burn Sepsis // Indian J. Med. Sci. — 1999. — 53, No. 5. — P. 216-219.
- Real R. Tables of Significant Values of Jaccard's Index of Similarity // Miscellania Zoologica. — 1999. — 22, No. 1. —29-40.
- HammerО., Harper D.A.T., Ryan P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. — 2001. — 4, No. 1. — P. 9.
- Ramette A. Multivariate analyses in microbial ecology // FEMS Microbiol. Ecol. — 2007. — 62, No. 2. — P. 142-160.
- Tegendre P., Tegendre T. Numerical Ecology. — Amsterdam: Elsevier Science BV, — 853 p.
- Андреева С.В., Бахарева Л.И., Шафикова Н.Э., Попова М.А., Угнивенко М.И. Особенности антибиотикочувствитель-ности этиологически значимой микрофлоры пациентов ожоговой реанимации / ТезисыXI междунар. конгресса МАКМАХ/ESCMID по антимикробной терапии. — 2009. — Т. 11, № 4. — С. 9-10.
- Андреева С.В., Бахарева Л.И., Валеева Д.М. Динамика антибиотикорезистентности у ведущих возбудителей ожоговых инфекций // Вестн. Челябинского гос. ун-та. — 2013. — № 7(298), Биология, Вып. 2. — С. 60-61.
- Похиленко В.Д., Перелыгин В.В. Бактериоцины: их биологическая роль и тенденции применения // Исследовано в России: Электронный науч. журн. — С. 164-198. —URL: http://zhurnal.ape.relarn.ru/articles/2011/016.pdf