В лесовосстановительной практике актуально cоздание стабильно функционирующих биоценозов, что способствует сохранению потенциала почв и лесного фонда. Данные почвеннозоологического анализа представляют интерес для определения динамики почвообразовательного процесса, общей направленности сукцессионных процессов, оценки возможной нагрузки на биоценоз и мер необходимого ухода. Особенно важна такая информация для заповедников и национальных парков. Стремясь к гармонии с окружающей природой, человек несет моральную ответственность за то, чтобы свободноживущие организмы сохранялись во всем многообразии форм, не подвергаясь массовому уничтожению в результате унификации и нарушения заселенных ими территорий. Сохранение потенциала лесного ландшафта, как одна из проблем окружающей среды, нуждается в правильной оценке возможной нагрузки и необходимого ухода. Оценка отклонения существующих и антропогенно преобразованных биоценозов от их естественного состояния (т.е. степени гемеробности) возможна по состоянию популяций педобионтов, которые являются не только участниками непосредственной переработки органических веществ, но и регуляторами микробиологической активности. Таким образом, синэкологические особенности педобионтов представляют интерес не только с зоологической точки зрения, но и для характеристики почвообразовательного процесса, направленности сукцессионных процесов в биогеоценозах.
Целью нашей работы являлось изучение биогеоценозов, образованных посадками сосновых культур в Баянаул-Каркаралинской группе реликтовых лесных оазисов для определения перспектив восстановления.
Лесовосстановительные работы в этом регионе связаны с трудностями поселения и произрастания сосны в аридном климате, на границе своего южного ареала распространения. Естественное возобновление сосны незначительно, особенно на вырубках и гарях, быстро зарастающих степными дерновинными злаками. В области гранитных интрузий на поверхности выветривающихся гранитов позиции сосны еще более ослаблены [1].
Закладка новых посадочных площадей способствует предупреждению деградации плодородных лесных почв в межгорных долинах и горных лесных черноземов. Немаловажно и значение лесовосстановления для предупреждения дальнейшей аридизации климата.
Характеристика района работы, материал и методика
Степная зона занимает центральную часть Казахского мелкосопочника. Основной растительный покров этой зоны состоит из разнотравно-злаково-ковыльных и типчаково-полынно-ковыльных ассоциаций. В разнотравно-ковыльной подзоне степной зоны сосна почти повсюду связана с гранитами, но встречается местами также на выходах кварцитов и метаморфических сланцев, а иногда и эффузивных горных пород. В типчаково-ковыльной подзоне степной зоны сосна связана исключительно лишь с гранитными низкогорьями и за их пределами отсутствует. Поскольку условия для обитания сосны недостаточно благоприятны, здесь она находится на пределе своего географического и экологического ареала [2].
В лесных оазисах Баянаульско-Каркаралинской группы наряду с сосновыми лесами встречаются своеобразные аридно-петрофитные сосновые редколесья; последние в некоторых оазисах даже превосходят по площади сосновые леса. Большая часть мелколиственных древостоев представлена производными типами леса.
Под сосновыми редколесьями развиты примитивные зачаточные или фрагментарные почвы, а под сосновыми лесами — бурые лесные петроморфные элювиированные, которые периодически по всему профилю подвергаются промыванию. Под сосняками каменисто-лишайниковыми почвы дресвянистые, сильноскелетные бурые лесные.
Островные боры Казахстана — остатки некогда сплошной полосы хвойных лесов, простиравшихся от Урала до Алтая. Мнение о реликтовом характере островных боров Казахстана и их фаунистических комплексов, древней связи с югом Сибири разделяют многие исследователи.
Казахские олигоценовые флоры генетически ближе к европейским, чем к сибирским. С миоцена по плиоцен растительность этой территории приобрела характер хвойного леса, сложились благоприятные условия для проникновения в мелкосопочник бореальных элементов флоры из северной части Западно-Сибирской равнины, с Алтая и с Южного Урала [3].
На протяжении последних 200 лет в результате рубок, лесных пожаров, а отчасти и выпаса скота площадь сосновых боров в Казахском мелкосопочнике уменьшилась, а в некоторых небольших массивах сосна была почти полностью истреблена.
Почвенно-зоологические исследования проведены в июне-августе под посадками сосны 7–8 лет и 25–30 лет. Количественные сборы выполнены по общепринятой методике [4]. Размер почвенных проб 50х50х40см. Почва разбиралась ручным способом, по слоям глубиной 10 см. Число проб при каждом обследовании было не менее 12. Общее количество взятых почвенных проб 80. Измерения температуры проводили коленчатыми термометрами Соренсена и лабораторными термометрами ТЛ-2. Влажность почвенных образцов определялась термовесовым методом. В работе использованы статистические методы и методы синэкологического анализа.
Результаты и их обсуждение
Посадки сосны занимают примерно 15 % площади оставшихся лесов. Около 9 % посадок имеют возраст 7–15 лет. Посадочные площади располагаются на склонах и их шлейфах, где сосновые насаждения соседствуют с кустарниковой типчаково-таволговой степью, частично смешиваясь с березняками.
Почвы под посадками дерново-лугово-лесные, аллювиальные, часто оглеенные. Слой почвы почти полностью покрывает поверхность, выходы гранитных глыб редки. Увлажнение местами неравномерное, происходит за счет притока влаги с более высоких частей гранитных низкогорий, выклинивания трещинных вод и атмосферных осадков.
Стационарные площади I были представлены сосновым древостоем. Средняя высота древостоев 0,8 м, возраст 7–8 лет. Подлесок редкий, из Rosa spinossima, Juniperus communis. Травяной ярус развит довольно хорошо, в его составе: Purola rotundifolia, Sedum acre, Pulsatilla patens, Lathurus pratensis. Проективное покрытие до 40–45 %.
Анализ почвенного разреза:
АО (0–2 см). Подстилка из хвои, корней травянистых растений;
AI (2–10 см). Темно-серый легкий суглинок, обильны корни деревьев, кустарников, трав;
АВ (10–20 см). Буровато-темно-серый средний суглинок, уплотненный, примесь гранитной дресвы. Светло-бурый, равномерно окрашенный легкий суглинок с обильной дресвой, пылеватый, много корневых остатков;
ВС (20–45 см). Бурый средний суглинок.
Стационарные площади II были представлены древостоем из сосны с незначительной примесью березы (9С1Б), сомкнутость крон 50–70 %. Высота древостоя 8–10 м, возраст 25–30 лет. Кустарниковый ярус развит умеренно, средняя высота 1–1,5 м, покрытие 50–60 %, состоит из Cotoneaster melanocarpa, Rosa spinossima, Spiraea hypericifolia, Rosa laxa, Ribes saxatile. Травяной покров густой (покрытие 60–80 %), ксеромезофильного характера, богатый по видовому составу: Filipendula vulgaris, Thalictrum foetidum, Fragaria viridis, Hieracium virosum, Libanotis sibirica, Galium verum, Phlomis tuberosa, Achillea nobilis и др.
Анализ почвенного разреза:
АО (0–5 см). Подстилка из хвои и кусочков коры, листьев трав, разложившаяся, с обильным мицелием;
АI (5–15 см). Средний суглинок темно-бурой окраски, задернованный, содержит корни деревьев;
AB (15–26 см). Более плотный, с выраженной ореховатой структурой, переплетен древесными корнями;
ВС (26–60 см). Слоистый, с прослойками тяжелого суглинка и супеси, местами мелкой дресвы.
Корней немного. На глубине 55–60 см ржавые пятна, глубже — сизые прослойки.
Для сезонной динамики температур этих стационарных площадей характерен максимум температуры в середине июня–июля (28,5 °С), минимумы отмечены в июне и конце августа. Среднепрофильные величины температуры с июня по август 18,2–18,4 °С. Разница максимальных температур в подстилке и горизонте АI достигает 12 градусов.
В режиме влажности отмечены значительные колебания. Минимальное содержание почвенной влаги наблюдается с конца июня по июль (максимум испарения), с некоторым повышением в августе, когда атмосферных осадков было больше. Среднепрофильное содержание влаги в почве в этот период 52,8 %.
При проведении почвенных раскопок на стационарных площадях I численность педобионтов была 58–70 экз/м2. Общее число видов 70–76. Доля хищных форм 54 %, доля фитосапрофагов 46 %. Обычны полужесткокрылые (Hemiptera). Преобладают жесткокрылые (Coleoptera) — 47 % от общего числа. Выявлено резкое доминирование некоторых видов, что, по-видимому, является результатом влияния дефицита влаги в почве. Среди Carabidae доминирует эвритопный вид Calathus melanocephalus (55 %), предпочитающий дерновые солоди. Из Elateridae особенно многочислен вид Selatosomus latus (38 %). Это вид, свойственный открытым пространствам. При переходе почв из степных условий к условиям леса его проникновение под полог, очевидно, объясняется хорошим прогреванием невысоких древостоев. Основная масса щелкунов на протяжении лета сосредоточена в подстилке. Преобладающие виды устойчивы к пониженным температурам.
Вертикальные перемещения личинок зависят от режима влажности и температуры почвы. Основное количество личинок (50 %) найдено на глубине 0–5 см.
Определение сезонной динамики температур на стационарных площадях II показало, что максимум приходится на июль (24,5 °С). Среднепрофильные температуры с июня по август 15,2–15,4 °С. Регулирующая роль в теплоотдаче и теплопоглощении хорошо сформировавшейся лесной подстилки сказывается в уменьшении разницы максимальных температур (4 °С) в горизонтах АО и АI. Содержание почвенной влаги несколько уменьшается в середине июля, когда спадает приток воды с низкогорий и незначительно количество атмосферных осадков. Среднепрофильное содержание влаги в почве в этот период 76,6 %.
В посадках сосны 25–30-летнего возраста на площадях II численность почвенного населения оказалась выше — 127,2 экз/м2. Общее число видов 120–136. В составе мезофауны преобладают жесткокрылые (Coleoptera) — 43 %, двукрылые (Diptera) и дождевые черви (Lumbricidae) составляют 20,1–28 % соответственно. Среди Coleoptera доминируют личинки долгоносика Poludrosus mollis. Возрастает численность Serica brunnea. Коэффициент обилия полужесткокрылых (Hemiptera) 1,08. Из двукрылых многочисленны личинки сем. Bibionidae (47 %). В подстилке сосредоточено 30 % беспозвоночных: слое 0–5см — 43 %, на глубине 5–15 см — 21 %. Таким образом, в условиях сосновых посадок II значительно изменилось соотношение трофических групп: доля фитосапрофагов и сапрофагов составляет две трети общей численности.
В 7–8-летних посадках сосны, расположенных на склонах и шлейфах гор на серых суглинистых почвах, червей не было обнаружено, что, возможно, является следствием частого дефицита влаги в почве. Однако в почвах посадочных площадей II, в условиях длительного сохранения влажности подстилки и почвенных горизонтов, обитают дождевые черви двух морфоэкологических типов: подстилочные — Dendrodrilus rubidus tenuis, Dendrobaena octaedra и собственно-почвенные — Aporrectodea caliginosa caliginosa, Eisenia nordenskioldi pallida. Общая численность червей 32 экз/м2, индекс плотности 12,21.
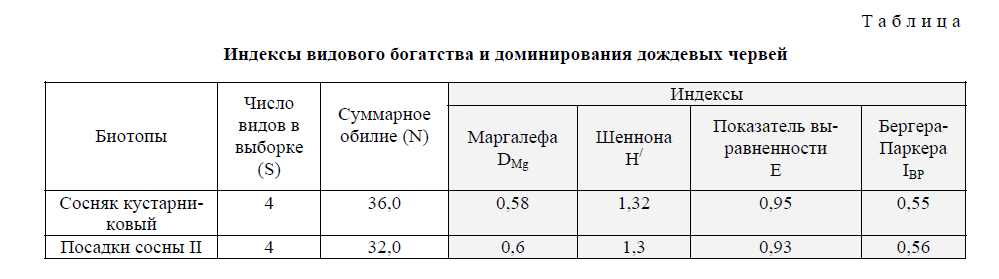
В естественных сухих и свежих сосновых лесах горного массива (сосняк каменистолишайниковый, сосняк травяно-костяничный, сосняк разнотравно-березовый, сосняк травяномшисто-папоротниковый) численность червей в почвенных пробах варьирует от 0,3 до 4 экз/м2, индекс плотности составляет 0,04–0,17. Только в сосняке кустарниковом, произрастающем в небольших ложбинах вдоль ручьев, также обнаружены черви четырех видов (рис.). Численность люмбрицид 36 экз/м2, соотношение видов 5:2:1:1 соответственно, индекс плотности составил 17,04. При сохраняющемся доминировании здесь D. octaedra другие виды имеют также соизмеримую численность, индекс Бергера-Паркера IBP=0,55 (табл.). Распределение обилия между видами стремится к равномерному, показатель выравненности E=0,95. Видовое богатство и разнообразие определялись по индексам Маргалефа DMg=0,58 и Шеннона H/ =1,32.
По данным Т.С.Перель, D. octaedra и E. nordenskioldi — доминирующий и сопутствующий виды восточных районов тайги [5]. D. octaedra и Dd. rubidus — основные виды сосняков-зеленомошников в средней и южной тайге. Следовательно, общий спектр видов в сосновых ценозах мелкосопочника близок таковому влажных сложных сосняков подзоны смешанных лесов. Примесь березы, осины, ольхи к сосновому древостою в этих лесах благоприятствует обитанию разных видов червей, увеличивая количество доступной органики в почве, уменьшая кислотность почв.
Рис. Соотношение жизненных форм дождевых червей в хвойных лесах Казахского мелкосопочника (в % от их общей численности)
В посадках сосны 25–30-летнего возраста доля почвенных видов в 2 раза ниже, чем в сосняке кустарниковом (соотношение 4:2:0,4:0,4). Однако согласно данным таблицы эти ценозы имеют близкие индексы видового богатства и разнообразия: DMg=0,60, H/ =1,3.
В почве стационарных площадей II доминирует D. оctaedra (19,8 экз/м2). В мелкосопочнике вид не встречается вне лесных ценозов, где обитает на буро-ложноподзоленных, горно-лесных, дерноволожноподзоленных, дерново-буро-ложноподзоленных почвах. Предпочитает почвы мощностью 30– 60 см и кислотностью рН 5,6–6,1. Наибольшая встречаемость его отмечена в сосняках с проточным увлажнением.
Индексы видового богатства и доминирования дождевых червей
Т а б л и ц а
Субдоминантным видом на стационарных площадях II является D. rubidus tenuis (12,2 экз/м2). Это космополитный вид монотипического рода Dendrodrilus, населяющий естественные природные ценозы. D. rubidus tenuis заселяет даже грубогумусную подстилку и способен быстро увеличивать свою численность при благоприятном режиме влажности. В неблагоприятный сухой период вид встречается в гниющей древесине, у корней, под камнями. Максимальная численность (84 % всех сборов) отмечалась в подстилке и верхнем почвенном слое естественных ценозов в мае, отмечено распределение по профилю почв на глубину до 15 см. Вертикальные перемещения, видимо, зависят от режима влажности и температуры.
Подобная сопряженность подстилочных видов D. оctaedra — D. rubidus tenuis характерна также для кустарниковых сосняков мелкосопочника. Во влажных лиственных лесах встречается близкое соотношение доминантных видов-гумусообразователей.
Как правило, в посадках наибольшая встречаемость дождевых червей зарегистрирована под комлем дерева, тогда как в подкроновых пространствах распределение лимитировано условиями мозаичной влажности. Такое распределение может быть отчасти объяснено тем, что большинство эпигейных видов предпочитает обитание в подстилке тех участков, где травянистый ярус выражен незначительно. Мозаичность распределения по биотопу может быть также связана с неравномерным увлажнением ценоза, где черви становятся своеобразным индикатором режима поступления влаги. Значительная доля молодых особей в весенний период (60–65 %) является критерием устойчивости этого компонента почвенной фауны. Кроме того, в условиях антропогенного преобразования ландшафта между видами A. caliginosa caliginosa — E. nordenskioldi pallida; A. caliginosa caliginosa — D. rubidus tenuis сложились отношения устойчивого сосуществования, отмеченные и в природных ценозах. Дальнейшее увеличение численности червей и приближение распределения к случайному (аналогично лесным сообществам) возможны при усилении задернения и мощности лесной подстилки, сопровождающихся более длительным удержанием влаги верхними почвенными горизонтами.
Близкая численность почвенных и подстилочных видов в естественных ценозах свидетельствует об активных процессах деструкции органических остатков и формировании устойчивого воздушного и водного режима почвенных горизонтов. Формирование подобного режима в лесопосадках показывает сближение их с естественными сосновыми лесами, что важно с точки зрения места закладки площадей и длительности планирования лесовосстановительных работ. Доминирующий бореальный вид D. octaedra является автохтоном, проникшим в мелкосопочник с бореальных элементми флоры из северной части Западно-Сибирской равнины, с Алтая и Южного Урала. Обнаружение многочисленных популяций таких видов в лесах и лесопосадках позволяет говорить о благоприятном состоянии интразональных реликтовых лесов Казахского мелкосопочника.
В целом в исследованных типах лесонасаждений сформировался комплекс подпологовой растительности и почвенной мезофауны, который отражает особенности температурного и гидрологического режима почвы каждой анализируемой площади.
Условия недостатка почвенной влаги в молодых сосняках, замедленность процессов образования подстилки и гумификации характеризуются присутствием большей частью луговых и полевых элементов почвенной мезофауны, обилием видов открытых пространств. В трофической структуре мезофауны представлены только хищные формы и фитосапрофаги. В структуре почвенного населения здесь преобладают насекомые, что является зональным признаком. Резко выраженная остепненность предполагает неустойчивое состояние ценоза.
Благоприятные условия сохранения почвенной влаги, более мощная подстилка и выровненный температурный режим в сосняках 25–30-летнего возраста благоприятствуют появлению лесных элементов мезофауны. Изменяется соотношение трофических групп: доля фитосапрофагов и сапрофагов составляет две трети общей численности. Наблюдается сходство фауны беспозвоночных с коренными мезофитными сосняками. Увеличение видового разнообразия педобионтов, доли сапрофагов и хищников в мезофауне почвы и в целом формирование трофической структуры, близкой к естественным сосновым ценозам, свидетельствуют о становлении более благоприятного экологического режима. На это же указывает успешное расселение дождевых червей. Преобладание фитофагов в герпетобии и хищных форм в почвенной мезофауне также указывает на начальные стадии сукцессионного процесса. Для дальнейшего развития ценоза как равновесной экосиcтемы необходимо соблюдение режима заповедного участка, не нарушаемого хозяйственной деятельностью.
Распространение червей в сосновые леса лимитировано условиями влажности, маломощными почвами, подстилаемыми близко залегающими гранитами или гранитной дресвой. Благоприятные для червей условия складываются в сосняке кустарниковом (36 экз/м), но и здесь доминируют подстилочные виды Dd. rubidus, D. octaedra в соотношении 5:2, распространение которых в почвах сосняков не ограничено условиями зимовки и связано с их морозоустойчивостью. Эти виды также способны осваивать и периодически засушливые, и переувлажненные биотопы.
Близкий видовой состав и соотношение видов червей в посадках сосны 25-летнего возраста (32 экз.м) обнаруживает сходство экологических режимов ценозов. Это позволяет сделать вывод о формировании лесного облика леопосадок, а доминирующие в них по численности подстилочные виды-вселенцы Dd. rubidus, D. octaedra рекомендовать для интродукции в молодые лесопосадки для активизации процессов гумусообразования. Встречающийся во влажных сосняках автохтонный вид Е. nordenskioldi хорошо зарекомендовал себя при интродукции в заболоченные ценозы для усиления процессов дернования в Джаныбеке [6]. Вид может быть аналогично расселен и в наших лесах, где обитание его, по нашим данным, благоприятствует повышению гумусообразования и дренированности почв [7].
Таким образом, при закладке посадочных площадей на дерново-лугово-лесных почвах в условиях Казахского мелкосопочника важен не только учет режима почвенной влажности, но и длительности процесса формирования биоразнообразия соснового ценоза.
Список литературы
- Горчаковский П.Л. Лесные оазисы Казахского мелкосопочника. — М.: Наука, 1987. — 159 с.
- Грибанов Л.Н. Сосновые леса Казахстана и биологические основы хозяйства в них. — Свердловск, 1965. — 54 с.
- Гвоздецкий Н.А., Николаев В.А. Казахстан. — М.: Мысль, 1971. — 296 с.
- Гиляров М.С. Методы почвенно-зоологических исследований. — М.: Наука, 1975. — 280 с.
- Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. — М.: Наука, 1979. — 272 с.
- Всеволодова-Перель Т.С. Формирование населения почвенных беспозвоночных (мезофауны) в лесных насаждениях комплексной полупустыни Прикаспия // Зоол. журн. — 2006. — Т. 85. — № 11. — С. 1327–1331.
- Абукенова В.С., Славченко Н.П. Региональные особенности почвенной мезофауны лесных ценозов Казахского мелкосопочника // Лесное почвоведение: итоги, проблемы, перспективы: Тезисы междунар. науч. конф. — Сыктывкар, — С. 96–97.