Известно, что от каждого органа или части тела отходят несколько лимфатических сосудов, которые на своём пути следования прерываются лимфатическими узлами. Количество лимфатических узлов у представителей различных видов млекопитающих широко варьирует. Так, их количество у свиньи составляет 190, у крупного рогатого ската 300, у мелкого рогатого скота 145 и у лошадей 8000. Средние размеры узлов колеблются от 0,9 до 10 см, реже до 20 см [1–7].
Впервые Н.М.Мырзахановым [8–11] установлено значение лимфатических узлов в транспорте жидкостей, который в опытах in vitro зарегистрировал ритмическое спонтанное сокращение лимфатических узлов тела (сомы) и внутренних органов, изучил иннервацию узла, влияние на них биологически активных веществ (норадреналин, ацетилхолин, гепарин, гистамин, АТФ) у ряда млекопитающих животных (крыса, овца, корова, свинья) и выдвинул концепцию о сосудисто-узловой системе транспорта лимфы в животном организме [11], которые нашли экспериментальное подтверждение в опытах [12], у Л.Э.Булебаевой [2] и М.Р.Хантурина [13].
Разработка научных проблем о генезе спонтанной ритмической сократительной функции лимфатических узлов, как и генез самой лимфатической системы, является мало изученным аспектом в учении о лимфатической системе. Согласно одной теории лимфатические сосуды произрастают из стенок вен, тогда как другая теория придерживается мнения о независимом от вен, мезенхимальном происхождении элементов лимфатической системы, а что касается вопроса о постнатальном становлении спонтанной ритмической сократительной активности лимфатических узлов и сосудов, вопрос остаётся открытым. Известно, что лимфатические узлы у плодов крупного рогатого скота и овец закладываются уже к третьему месяцу утробной жизни [3,6,7]. С возрастом узлы уплотняются и утолщаются их соединительно-тканные и гладкомышечные элементы. Внутренняя структура лимфатических узлов зависит от их локализации: в области сомы, в брюшной и грудной полостях [5,14]. По данным [14] ягнята рождаются с функционально сформировавшейся симпатической иннервацией лимфатических узлов, что чётко выявлялось при анализе содержания катехоламинов в узлах в различные сроки постнатального периода.
Согласно исследованиям последних лет (1) лимфатическая система млекопитающих представлена прелимфатическими и лимфатическими капиллярами, внутриорганными лимфатическими сосудами, отводящими афферентными лимфатическими сосудами, лимфатическими узлами, грудным и правым лимфатическими протоками.
К настоящему времени хорошо изучено тонкое структурно-топографическое строение и иннервация аппарата лимфообращения, установлены основные функции (дренажная, иммунологическая, белоктранспортная и.т.п.) лимфатической системы, которые обеспечивают сохранение эндоэкологического гомеостаза внутренних сред организма.
Функциями лимфатических узлов являются: лимфоцитопоэз, барьерно-фильтрационная, транспортная, иммунопоэз, обменная, депонирование (в частности, витаминов А и В), внутренней секреции (гепарин), разрушение эритроцитов. Естественно предположить, что в выполнении перечисленных выше функций лимфатических узлов немаловажное значение имеет изучение становления спонтанной ритмической сократительной функции лимфатических узлов, в частности, в постнатальном онтогенезе.
Материалы и методика исследований
Объектом исследования служили изолированные, одиночные соматические (подчелюстный, подмышечный, паховый) и висцеральные (сердечный, кишечный, брыжеечный, почечный, печеночный) лимфатические узлы лабораторных крыс, котят, сурков и телят.
Изометрическое напряжение препаратов регистрировали при помощи высокочувствительных механотронов типа 6 МХ-1Б на чернильно-пишущем самописце Н-327–5. Термостатирование производили с помощью ультратермостата УТ-15 с точностью не ниже 0,2 °С [4, 11].
Спонтанные ритмические сокращения лимфатических узлов производили на 1, 5, 10, 15, 20, 30, 60 и 90-й дни после рождения ягнят. Анализ лимфонодуллограммы производили по общей картине кривой относительно изометрической линий, а также по частоте и амплитуде сокращений узла, включающей 5 записей по одной минуте в течение 5 минут. Кроме того, с помощью курвиметра определяли продолжительность кривой за 5 минут, которая служила характеристикой общего уровня биоэлектрической активности узла [15]. Всего произведено более 3800 регистраций лимфонодуллограмм.
Результаты исследований
Результаты исследований показали, что характер спонтанной ритмической сократительной деятельности лимфатических узлов имеют чёткие отличия, в зависимости от сроков постнатального онтогенеза органа (табл.1)
Т а б л и ц а 1
Частота спонтанных сокращений лимфатических узлов у крысят в онтогенезе
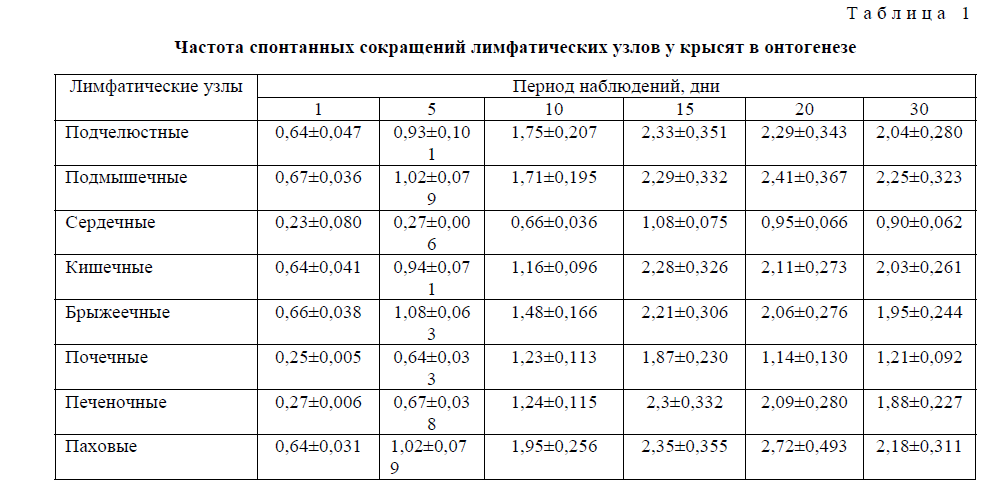
К моменту рождения спонтанная сократительная активность всех исследованных лимфатических узлов низка или практически отсутствует, а наблюдающиеся сокращения нерегулярны и спонтанны, без определённой закономерности. На 5-й день после рождения отмечается некоторое повышение сократительной деятельности узлов. Однако всё ещё не обнаруживается регулярность всех компонентов (частотной и амплитудной) сократительной деятельности лимфатических узлов, хотя общий уровень биоэлектрической активности заметно повышается.
На 10-й день постнатальной жизни выявляется характерная для органов специфичность кривой лимфонодуллограммы, сокращения становятся ритмическими, и выявляется частотно-амплитудная специфичность сократительной деятельности лимфатических узлов. Начиная с 15-го и по 30-й день обнаруживается характерная для взрослых крыс картина спонтанной ритмической деятельности лимфатических узлов. Установлено, что во все периоды постнатальной жизни частотно-амплитудные характеристики сократительной деятельности узлов меняются однонаправленно, т.е. синергично. Наиболее высокие частота и амплитуда сокращения узлов выявлены на 15-й день, с последующим сохранением указанных характеристик на 20–30-е сутки, за исключением почечных лимфатическим узлов, частотно-амплитудная характеристика снизилась в среднем на 40 % на 20 и 30-е сутки. Следует заметить, что сократительная деятельность лимфатических узлов такого органа, как сердце, обладающего автоматизмом, была низкой по сравнению с сокращением других висцеральных и соматических узлов. Это, по-видимому, связано с тем, что в циркуляции лимфы и внеклеточной жидкости значительная роль принадлежит мышечному микронасосу сердца (Косицкий,1975), который работает постоянно.
Характер спонтанной ритмической сократительной деятельности лимфатических узлов котят показал чёткие отличия, в зависимости от сроков постнатального онтогенеза органа (табл.2)
Т а б л и ц а 2
Частота спонтанных сокращений лимфатических узлов у котят в онтогенезе
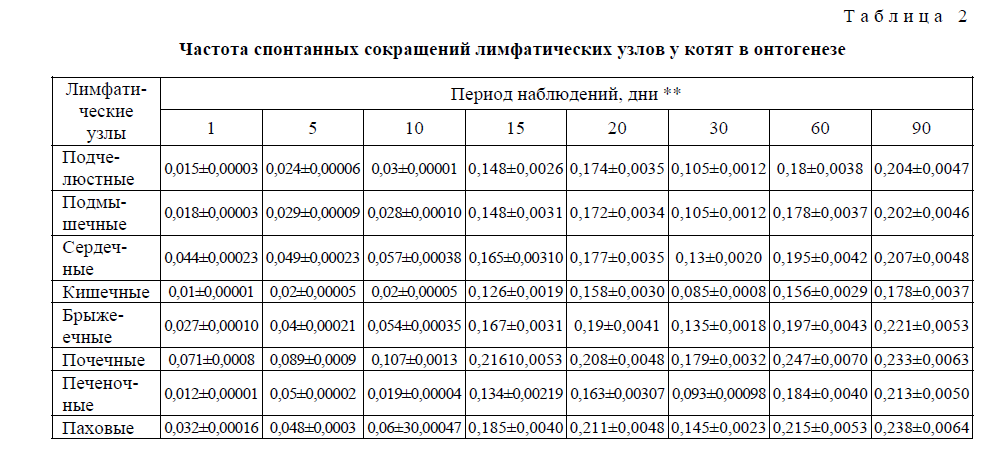
Направление формирования частотной и амплитудной характеристик, составляющих спонтанную ритмическую сократительную активность лимфатических узлов в ходе постнатального развития котят имеет аналогичную форму. Как видно из таблицы 2, к моменту рождения и в последующие 50– 10 дней, а для отдельных лимфоузлов и до 15 (сердечные, паховые) дней спонтанная сократительная активность всех исследованных лимфатических узлов низка или практически отсутствует. Наблюдающиеся сокращения нерегулярны и спонтанны, без определённой закономерности. На 15-й день после рождения отмечается достаточно высокая активация сократительной деятельности узлов, за исключением сердечных (по амплитуде) и паховых (по амплитуде и частоте) лимфатических узлов. Обнаруживается регулярность всех компонентов (частотной и амплитудной) сократительной деятельности лимфатических узлов с одновременным повышением общего уровня биоэлектрической активности. На 30-й день постнатальной жизни, а для кишечного лимфатического узла на 20-й день (по амплитуде) выявляется снижение характерной для органов специфической кривой лимфонодуллограммы с последующим повышением на 60 и 90-й дни постнатального развития.
Исследования спонтанной ритмической сократительной деятельности лимфатических узлов сусликов и телят показали чёткие отличия, в зависимости от сроков постнатального онтогенеза органа (табл. 3–4).
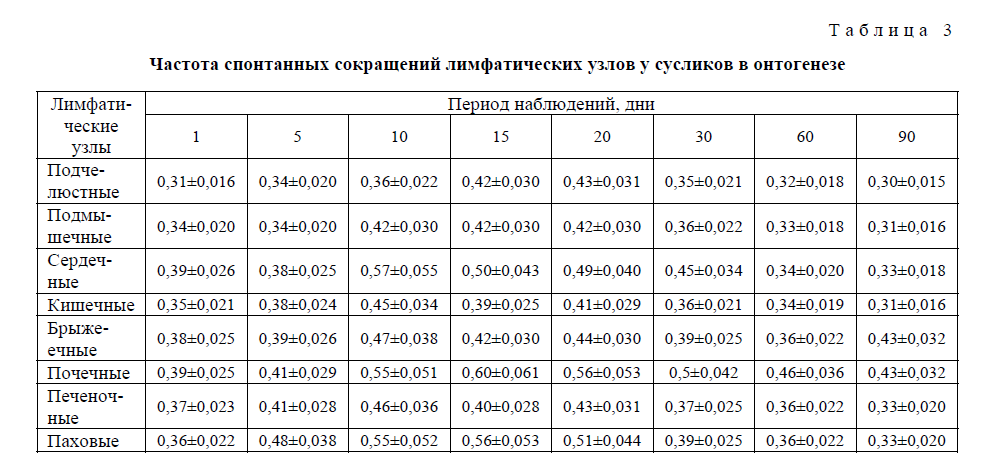
Т а б л и ц а 3
Частота спонтанных сокращений лимфатических узлов у сусликов в онтогенезе
Т а б л и ц а 4
Частота спонтанных сокращений лимфатических узлов у телят в онтогенезе
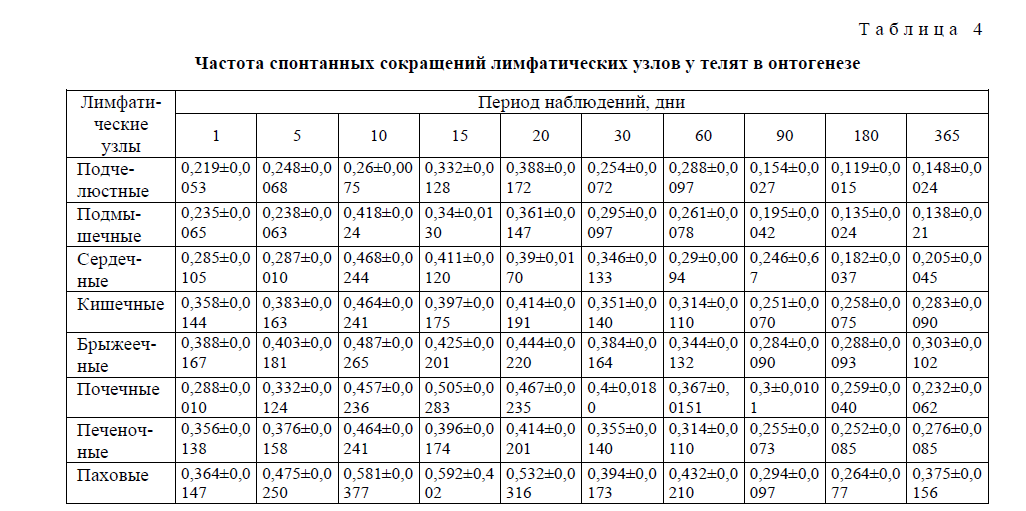
Направление формирования частотной и амплитудной характеристик, составляющих спонтанную ритмическую сократительную активность лимфатических узлов в ходе поснатального развития сусликов, имеет аналогичную форму. К моменту рождения и последующие 50–10 дней, а для отдельных лимфоузлов и до 15 (брыжеечные) дней спонтанная сократительная активность всех исследованных лимфатических узлов низка или практически отсутствует, а наблюдающиеся сокращения нерегулярны и спонтанны, без определённой закономерности. На 15-й день после рождения отмечается достаточно высокая активация сократительной деятельности почечных, паховых, сердечных, подчелюстных, подмышечных, печеночных и кишечных лимфатических, за исключением брыжеечных (по частоте), лимфатических узлов. Обнаруживается регулярность всех компонентов (частотной и амплитудной) сократительной деятельности лимфатических узлов с одновременным повышением общего уровня биоэлектрической активности. На 15-й день постнатальной жизни, а для брыжеечного лимфатического узла — на 20-й (по частоте и амплитуде), выявляется характерная для органов специфическая кривая лимфонодуллограммы с последующим повышением на 60 и 90-й дни постнатального развития. В эти же промежутки времени обнаруживается характерная для взрослых телят картина спонтанной ритмической деятельности лимфатических узлов. Установлено, что в целом в постнатальном периоде жизни телят частотно-амплитудные характеристики сократительной деятельности узлов меняются однонаправленно, т.е. синергично, с небольшими хронологическими отличиями между узлами.
Установлено, что у сусликов и телят происходит активация спонтанной сократительной деятельности лимфатических узлов между 15 и 20-м, а также 60 и 90-м днями после рождения, с некоторым снижением этой активности на 30-й день постнатального развития. В эти же промежутки времени обнаруживается повышение общего уровня биоэлектрической активности лимфатических узлов.
Таким образом, нами впервые установлены видовые особенности формирования спонтанной сократительной деятельности лимфатических узлов тела и внутренних органов, которые осуществляются в виде двуфазной хронолого-пространственной организации моторной деятельности лимфатических узлов.
Выводы
- У всех исследованных млекопитающих спонтанная ритмическая сократительная активность лимфатических узлов сомы и висцеральных органов в первые дни после рождения практически не выявляется.
- Активация спонтанной сократительной деятельности лимфатических узлов тела и внутренних органов происходит дважды — между 15 и 20-м, а также 60 и 90-м днями после рождения, с падением этой активности на 30-й день постнатального онтогенеза.
Список литературы
- Бородин Ю.И. Функциональная морфология. — Новосибирск, 1992. — С. 12–17.
- Булекбаева Л.Э. Корковая лимбическая система и регуляция лимфотока. — Алматы, 1976. — 286 с.
- Григорьев В.С. Возрастные изменения ретикулярной ткани и клеточного состава брыжеечных лимфатических узлов у плодов крупного рогатого скота. — Уч. зап. Казанского ветер. института. — Казань, 1975. — Т.120. — С. 170–174.
- Ерофеев Н.П., Мандрыко Е.С. Модификация системы для исследования мышечных препаратов // Физиол. журн. ШР, 1975. — Т.61. — № 8. — С.
- Игнатьев Н.Г. О функциональной активности симпатической активации некоторых лимфатических узлов в различные фазы постнатального периода развития. — Уч. зап. Казанского ветер. института. — Казань, — Т.118. — С. 235– 237.
- Иосифов Г.М. Лимфатическая система овцы. — Орджоникидзе, 1942. — 96 с.
- Кривский И.Л. Топография и особенности строения тазовых и перикортальных лимфатическиз узлов собаки. // Материалы конф., — М., 1965. — С. 45–56.
- Мырзаханов Н.М. Влияние различных рационов на лимфообращение // Тр. междунар. конф. по лимфообращению. —Новосибирск, 1992. — С. 87.
- Мырзаханов Н.М. Экспериментальное изучение сократительной активности лимфатических узлов // Вестник НАН РК.— № 4. — С. 46–53.
- Мырзаханов Н.М. Роль лимфатических узлов и сосудов в продвижении лимфы // Вестник НАН РК. — № — С. 70–76.
- Мырзаханов Н.М. Функциональные особенности лимфообращения сельскохозяйственных животных: Автореф. дис... д-ра биол. наук.— Алматы, 1995. — 42с.
- Орлова Р.С. Лимфатические сосуды. — Л., 1982. — 362 с.
- Славсизон Л.Д. К вопросу о регионарных особенностях лимфатических узлов у кролика. — Донбасс, — В. 1. —С. 44–46.
- Собакин М.А. Экспериментальная методика электрографического исследования моторной деятельности желудка при пищеварении // Биол. эксп. биол. и мед. 1953. — Т. зв. — В. 9 — С. 76–79.
- Хантурин М.Р. Сравнительная характеристика и механизмы сократительной деятельности лимфатических сосудов: Автореф. дис... д-ра биол. наук. — Алматы,