В данной работе рассмотрена организация клетки как системы внутриклеточных микроструктур и метаболических процессов по сложности не уступает организации организма человека как системы органов, тканей и клеток. Однако изучать жизнедеятельность клетки по ее биоритмам проще, потому что процессы на клеточном уровне происходят быстрее. Биоритмы клетки по сравнению с биоритмами организма имеют более короткие периоды, постоянные времени обратных связей в контурах регуляции, а переходные процессы смены режимов функционирования короче, что важно для постановки серий опытов. Изучение временной динамики процессов в клетке может использоваться для диагностики заболеваний организма человека и их лечения. Механизмы развития болезней связаны с нарушением обмена веществ и микроструктуры клеток. Судить о них и оценивать эффективность используемого метода лечения возможно по виду, степени и характеру изменений биоритмов клетки, т. е. возникающих рассогласований во времени внутриклеточных процессов.
Введение. Сегодня накопленные хронобиологией факты позволили наполнить конкретным содержанием многие предположения о происхождении и эволюции жизни, понять и обьяснить явление адаптации организмов к меняющейся внешней среде, выявить закономерности взаимосвязанной деятельности систем внутренней среды организма, определить принципиально новые пути в понимании нормы и патологии. Хронобиология открыла неизвестные ранее подходы к диагностике, лечению и профилактике болезней. Ибо биологические объекты, как открытая термодинамическая система, в своем развитии впитали в себя все закономерности ритмически колеблющегося внешнего мира и взаимодействуют с ним при помощи этих колебаний.
До настоящего времени многие понятия в биологии и медицине исходят из стационарности физиологических процессов, осуществляющихся на одном (нормальном, нативном) уровне или изменяющихся линейно. Формирование биолога или врача в ВУЗ-е часто идёт на основе представлений о физиологических константах, которых никогда не бывает в действительности. И это факт,что в классической биологии общеприняты незыблемые постоянства тех или иных функциональных проявлений, а всеобщность ритмического феномена в физиологических процессах и сейчас многими игнорируется.
Междутем только через периодические колебания всех жизненных функций - от энергетических и молекулярных превращений до изменения численности популяций, - связанные с колебательными контурами ближней и дальней внешней средой, осуществляется нормальная жизнедеятельность [1-3].
Поэтому на современном этапе развития хронобиологии любые исследования, заполняющие, как кирпичиками, ещё пустующие ниши нового знания, отражают передовые рубежи естествознания и включаются в создание будущей общей теории биологии. Подтвержением этому являются ряд открытии ученых в области физики, физиологии и медицине.
Так, лауреатами Нобелевской премии в области физики в 2017 году стали группа ученых за открытие гравитационных волн, а в области физиологии и медицины лауреатами Нобелевской премии стали ряд ученых (Холл, Росбали, Янг) за цикл работ по теме: «Изучение механизмов суточной (циркадианной) динамики организма».
Сегодня хронобиология может охарактеризовать периодические колебания, пожалуй, всех органов, всех систем организма. Накоплено огромное количество данных о суточных и других ритмах органов пищеварения и мочевыделения, дыхательной и сердечно- сосудистой систем, головного мозга и т.д. [ 4-10].
Как ни странно, но вне достаточного внимания ученых, занимающихся проблемами хронобиологии, оказались биологически активные точки (БАТ) кожи, хотя, казалось бы, именно они должны были бы в первую очередь стать объектом хронобиологических исследований, актуальность которых несомненна. Такую же актуальность имеет вырос временной организации гормонов при различных функциональных состояниях организма.
Актуальность исследований в этих направлениях несомненна, ибо возможно получать значительную информацию не только о самом нарушенном процессе, но и дополнить имеющиеся представления о биоактивных точках и гормонах как о биоритмологических показателях состояния внутренних систем организма.
Целью исследования, является установление хроноструктурных параметров суточной (циркадианной) динамики электрических свойств биоактивных точек коже и гормонов надпочечников у животных в норме и при стрессе.
Материалы и методы. В качестве объектов исследования использовали самцов лабораторных белых крыс с массой тела 250 - 270 гр. И кроликов породы шиншилла, обоегопола, массой 1000-1300 гр.
Были сформированы 4 группы животных, каждая из которых была подразделена на 3 подгруппы. В эксперименте были задействованы 90 крыс и 15 кроликов. Параметры статических электрических потенциалов (СЭП) измеряли на коже в области расположения аурикулярных БАТ №№ 1,2 и 3, расположенных симметрично на обоих ушных раковинах и проецирующих миндалины [11].
Экспериментальную модель ангины у кроликов вызывали по известной методике [12] Измерение СЭП производили с помощью известной методики [13-15]
Гипокинетический стресс моделировали помещением крыс в клетки - пеналы. Размер клеток составлял 7х 12 см, сверху натягивалась металлическая сетка.
Гормоны надпочечников определяли при помощи флюорометрического метода [16,17] и спектрофотометра Hitachi - 650- 60.
Все экспериментальные данные подвергались статистической обработка с нахождением критерия достоверности различий, а хроноструктурные параметры суточных биоритмов выявляли и анализировали с помощью «Косинор» анализа[18-20].
Результаты и их обсуждение. Результаты исследований суточной динамики статических электрических потенциалов (СЭП) биоактивных точек (БАТ) кроликов в норме и при стрессе представлены на рисунок 1 и 2.

|
№№ БАТ |
Среднесуточная величина (мезор) (доверит. интервал) |
Амплитуда (А) ± от М (доверит. интервал) |
Акрофаза в ч и мин (доверит. интервал) |
Период (Р), ч | |
|---|---|---|---|---|
|
1 |
2 |
3 |
4 |
5 |
|
N° 1 |
50,8 ± 1,9 (44,9 ± 0,8 ÷ 56,7 ± 2,5) |
5,9 (5,9 ÷ 6,0) |
00 ч 30 мин (23 ч 24 мин ÷ 01 ч 30 мин) |
24 |
|
№ 1' |
50,6 ±1,8 (43,3 ± 1,0 ÷ 60,1 ± 2,4) |
8,4 (7,3 ÷ 9,5) |
00 ч 28 мин (23 ч 20 мин ÷01 ч 26 мин) |
24 |
|
№2 |
52,5 ±2,2 (46,4 ±2,0 ÷ 60,0 ± 2,9) |
I 6,8 (6,1 ÷7,5) |
00 ч 18 мин (23 ч 30 мин ÷ 01 ч 06 мин) |
24 |
|
№ 2' |
52,9 ±2,1 (45,5 ± 2,2 ÷ 64,0 ± 2,5) |
9,2 (7,4 ÷ 11,1) |
00 ч 22 мин (23 ч 34 мин ÷ 01 ч 10 мин) |
24 |
|
№3 |
56,1 ±2,0 (50,8 ± 1,2 ÷ 66,8 ± 2,0) |
8,0 (5,2 ÷ 10,8) |
23 ч 42 мин (23 ч 30 мин ÷ 00 ч 00 мин) |
24 |
|
№3' |
54,7 ±2,0 (46,0±2,7 ÷ 67,1 ± 1,9) |
10,1 (7,8 ÷12,4) |
23 ч 40 мин (23 ч 30 мин ÷ 00 ч 06 мин) |
24 |
Таблица 1 - Косинор-анализ суточного ритма статических электрических потенциалов (в мВ) аурикулярных биоактивных точек: №№1,2,3 И 1'2'3' кожи кроликов в норме
Судя по усредненным показателям (рисунок 1), уровень СЭП всех измеренных БАТ в норме синхронно колебался по типу одновершинной кривой с максимумом в 22- 24 ч. и сравнительно низкими значениями в утренне - дневные часы. Так, величины СЭП БАТ № 1 (правая) в течении суток варьировали от 44,9± 0,8 до 56,8 ± 2,5 мВ, БАТ № 1′(левая )- от 43,3 ± 1,0 до 60,1 ± 2,4 мВ, БАТ № 2 – от 46,4± 2,0 до 60,0 ± 2,9 мВ, БАТ № 2' – от 45,5± 2,2 до 64,0 ± 2,5 мВ, БАТ № 3 – от 50,8 ± 1,2 до 66,8 ± 2,0 мВ и БАТ №3 ' 46,9 ± 2,7 до 67,1 ± 1,9 мВ. Полученные данные отчетливо показали, что форма волны и уровни фиксированных на шкале времени величин СЭП при измерении в течение 2,5 суток были одинаковыми, а левые части суточных кривых были аналогичны таковой первый половина треьих суток. При этом величины СЭП симметричных БАТ левого и правого уха совпадали как по направленности суточных изменений, так и по абсолютных значениям в фиксированные часы суток.
Таким образом, уже визуальный анализ динамики СЭП аурикулярных БАТ кожи на протяжении более 2-х суток даёт основание считать, что выявленные колебания СЭП являются четким суточным ритмом с наивысшими показателями в первый половине ночи.
Достаточно веским подтверждением наличие хорошо выраженного суточного ритма СЭП доказывают расчёты и анализы, проведенные по программе – методике «Косинор» (таблица 1): акрофаза во всех случаях располагалась в пределах полуночи; мезор СЭП в симметричных БАТ совпадали, а его значения в несимметричных БАТ были близки; в большей степени различались амплитуды всегда превышали значения правой.
колебаний СЭП, которые в БАТ левой ушной раковины
Таблица 2 - Косинор -анализ суточного ритма статических электрических потенциалов (в мВ) аурикулярных биоактивных точек №№ 1, 2, З и 1", 2 3' кожи кроликов при стрессе
|
№№ БАТ |
Среднесуточная величина (мезор) (доверит. интервал) |
Амплитуда (А) ± от М (доверит. интервал) |
Акрофаза в ч и мин (доверит. интервал) |
Период (Р), ч | |
|
1 |
2 |
3 |
4 |
5 |
|
№ 1 |
69.5 ±2,5 (60.8 ±2,3÷79,1 ±2,8) |
9,1 (8,7 ÷ 9,6) |
02 ч 42 мин (00ч 24мин ÷ 04ч 54мин) |
24 |
|
№ 1 |
69,5 ±2,5 (60,8±2,3÷79,1 ± 2,8) |
7,2 (6,4÷ 8,0) |
03 ч 30 мин (01ч 42мин ÷ 05ч 24мин) 11 ч 30 мин (09ч 42мин ÷ 13ч24мин) 19 ч 30 мин 17 ч 42 мин÷ 21 ч 24 мин) |
08 |
|
№ 1’ |
76,5 ±2,6 (66,4 ± 2,9 ÷ 85,9 ±2,4) |
9,8 (9,2 ÷ 10,3) |
02 ч 38 мин (00ч ЗОмин ÷ 04ч 50мин) |
24 |
|
№ 1’ |
76,5 ±2,6 (66,4 ± 2,9÷ 85,9 ±2,4) |
7,6 (6,7÷ 8,5) |
03 ч 26 мин (01Ч36ИН÷ 05Ч 20МИН) 11 ч 26 мин (09 ч 36 мин ÷13 ч 20 мин) 19 ч 26 мин |
08 |
|
№ 2 |
68,5 ±2,7 (57,1 ±2,8÷ 78,8 ±2,5) |
10,8 (10,3 ÷11,4) |
01 ч 48 мин (21ч12мин÷ 05ч 54мин) |
24 |
|
№ 2’ |
68,5 ±2,7 (57,1 ±2,8÷ 78,8±2.5) |
9,7 (9,1 ÷10,3) |
03 ч 18мин (02ч З6 мин ÷ 04ч Обмин) 11 ч 18 мин (10 ч 36 мин ÷12 ч 06 мин) 19 ч 18 мин |
08 |
|
№ 2’ |
77,5 ±2,5 (69,3 ± 2,4 ÷ 89,9 ±2,4) |
10,3 (8,2 ÷ 12,4) |
01 ч 56 мин (21ч 18мин ÷ 05ч 58мин) |
24 |
Таким образом, у здоровых кроликов выявлен хорошо выраженный синфазный одновершинный суточный ритм СЭП БАТ обеих ушных раковин (Р ≤0,001) изначения хроноструктурных параметров биоритмов (период, амплитуда, мезор, акрофаза), так с акрофазой приходящейся на полночь и минимумом в утреннее - дневное время, амплитудой от 5,9 до 10,1 мВ, мезором от 50,6 до 56,1 мВ и периодом в 24 час.
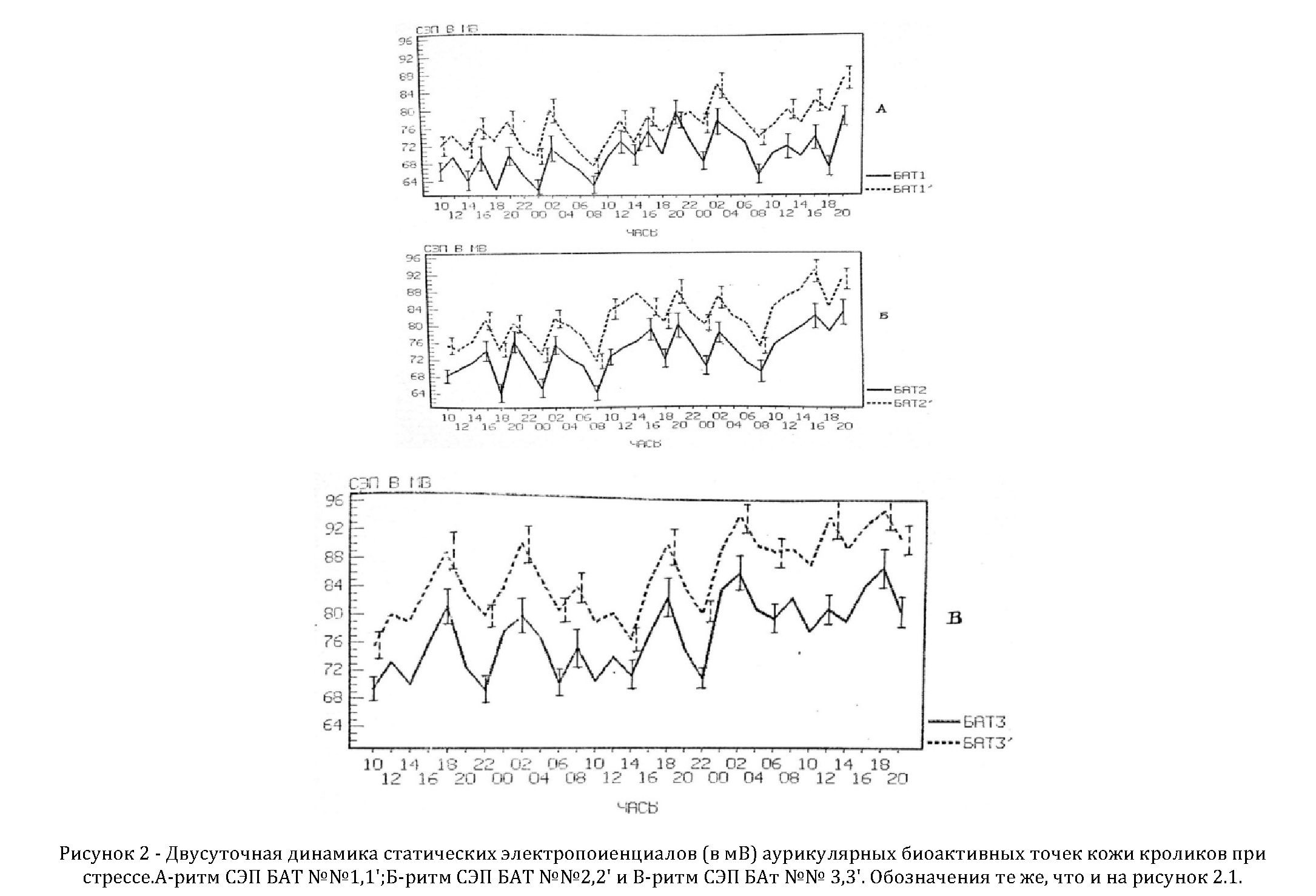
Также нами были изучены и установлены суточные (циркадианные) ритмы СЭП БАТ кроликов при стрессе (ангине).
При стрессе установлены исчезновение одновершинности суточных колебаний СЭП, они приобретали полифазный характер. Обращало внимание существенное увеличение разброса абсолютных значений СЭП и возрастание в 1,3 раза вольтажа БАТ (рисунок 2). Кроме этого, была обмечена потеря симметричности показателей СЭП правой и левой ушной раковины, столь характерной для суточного ритма СЭП у здоровых кроликов, также было показано, что величины СЭП в ночное время снижались и повышались днем.
Таблица 2 (Продолжение)
|
№№ БАТ |
Среднесуточная величина (мезор) (доверит. интервал) |
Амплитуда (А) ± от М (доверит. интервал) |
Акрофаза в ч и мин (доверит. интервал) |
Период (Р), ч | |
|
1 |
2 |
3 |
4 |
5 |
|
№2 |
77,5 ±2,5 (69.3 ±2,4÷ 89.9 ±2.4) |
9,1 (8,1 ÷ 1 0,1) |
03 ч 26 мин (02ч 40ин ÷04ч 12мин) 11 ч 26 мин (10 ч 40 мин ÷12ч 12 мин) 19 ч 26 мин |
08 |
|
№3 |
68.8 ±2,4 (55,1 ±2,6 ÷ 78.4 ±2.2) |
11.6 (9,6 ÷ 13,7) |
21 ч 12 мин (17Ч36МИН÷ 23ч 54МИН) |
24 |
|
№3 |
68.8 ±2.4 (55.1 ± 2.6 ÷78.4 ±2.2) |
10,2 (9,0 ÷11,4) |
03 ч 12мин (01ч54мин ÷04ч З6мин) 11 ч 12 мин (09 ч 54 мин ÷12 ч 36 мин) 19 ч 12 мин (17 ч 54 мин 4- 20 ч 36 мин) |
08 |
|
№3’ |
78,4 ±2.7 (67,4 ±2.9 ÷89.3 ±2.9) |
10.9 (10.9÷11,0) |
21 ч 14 мин (17ч38мин 4- 23ч 56мин) |
24 |
|
№3’ |
78,4 ±2,7 (67.4 ±2.9 ÷89.3 ±2.9) |
9,5 (8.5 ÷10,5) |
03 ч16мин (01ч 58мин 4- 04ч 40мин) 11 ч 16 мин (09 ч 58 мин 4- 12 ч 40 мин) 19 ч 16 мин |
08 |
Расчеты и анализы по программе «Косинор» позволили найти, что акрофазы 24-часовой периодичности колебаний СЭП в БАТ № 1 и 2 сместились в сторону ночных часов и регистрировались в 02 час 42 мин. в первом случае и 01 час 48 мин. – во втором (таблица 2). Мезор СЭП у кроликов при стрессе в БАТ № 1 и 2 возрос почти на 35 %, при таком же усилении амплитуд. Полностью были установлены значения хроноструктурных параметров биоритмов при стрессе (таблица 2).
Таким образом, моделирование у кроликов состояния стресса, которое возникало при развитии у животных ангины, приводило к расслоению суточного ритма СЭП БАТ на непосредственно 24-часовой и 08-часовой периоды. Этот 8-часовой период на протяжении суток формировал три акрофазы, что отражало резкое изменение суточного ритма СЭП при стрессе.
Все эти изменения ритма СЭП можно отценить однозначно: воспалительный процесс, являясь мощным стрессовым фактором, дезорганизует облаженную суточную биоритмику в БАТ, что отражает возникновение выраженного внутренного десинхроноза.
Таблица 3 - Суточная динамика адреналина в плазме крови крыс в норме и при стрессе
|
Время суток, ч |
Адреналин, нг/мл |
|
|
Контроль, n = 21 |
опыт, n =21 |
|
|
10 |
0,054±0,017 |
0,019±0,005 |
|
11 |
0,045±0,002 |
0,030±0,005 |
|
12 |
0,065±0,002 |
0,067±0,012 |
|
13 |
0,007±0,001 |
0,032±0,012 |
|
14 |
0,045±0,009 |
0,029±0,006 |
|
15 |
0,034±0,010 |
0,083±0,029 |
|
16 |
0,045±0,002 |
0,093±0,027 |
|
17 |
0,035±0,006 |
0,027±0,003 |
|
18 |
0,085±0,006 |
0,027±0,002 |
|
19 |
0,060±0,015 |
0,019±0,005 |
|
20 |
0,045±0,006 |
0,049±0,011 |
|
21 |
0,050±0,004 |
0,030±0,005 |
|
22 |
0,051±0,001 |
0,040±0,006 |
|
23 |
0,070±0,001 |
0,015±0,004 |
|
24 |
0,065±0,002 |
0,033±0,007 |
|
01 |
0,040±0,008 |
0,047±0,013 |
|
02 |
0,085±0,002 |
0,043±0,002 |
|
03 |
0,032±0,002 |
0,018±0,004 |
|
04 |
0,035±0,002 |
0,037±0,004 |
|
05 |
0,030±0,004 |
0,020±0,001 |
|
06 |
0,045±0,009 |
0,038±0,001 |
|
07 |
0,045±0,002 |
0,020±0,001 |
|
08 |
0,014±0,002 |
0,027±0,002 |
|
09 |
0,035±0,002 |
0,030±0,001 |
Судя по усредненным показателям (таблице 3), уровень адреналина в контрольных крыс в течение суток подвергался колебательным изменениям в максимумами и минимумами в разные часы суток.
Среднесуточное содержание адреналина в плазме крови интактных крыс составляет около 0,047 нг/мл (см таблицу 3). Разброс значений в течение суток значителен: индивидуальные значения варьируют от 0,005 нг/ мл до 0,1 нг/мл.
Усредненные значения концентрации адреналина в норме варьировалиот 0,007±0,001 нг/ мл до 0,085 ± 0,006 нг/ мл. Минимальные показатели адреналина фиксировались в 13 час, максимальные в 18 час и 02 час.А усредненные показатели адреналина у подопытных (при стрессе) животных в течение суток варьировали от 0,015± 0,004 нг/ мл до 0,093±0,027 нг/мл и значительно отличали от таковых в норме как по конфигурации биоритмов, так и по концентрации гормона.
Концентрация норадреналина, также, как и адреналина, подвержены суточным колебаниям как в норме, так и при стрессе (таблица 4).У контрольных групп животных содержание норадреналина в течение суток варьирует от 0,025± 0,002 нг/мл до 0,250±0,019 нг/мл. Общее содержание норадреналина, как адреналина у опытных групп животных меньше, чем у интактныхгрупп крыс. Максимальные значения норадреналина у животных в норме приходятся на 03 часа ночи, минимальные показатели определены в 17 часов, при стресс максимальные значения концентрации норадреналина приходятся на 10 – 12 часов, а минимальные на 03 и 06 часов.
Таблица 4 - Суточная динамика нороадреналина в плазме крови крыс в норме и при стрессе
|
Время суток, ч |
Норадреналин, нг/мл |
|
|
Контроль, n = 21 |
Опыт, n = 21 |
|
|
10 |
0.130±0.026 |
0.127±0.019 |
|
11 |
0,135±0,029 |
0,083±0,008 |
|
12 |
0.105±0,036 |
0,167±0,017 |
|
13 |
0,120±0,030 |
0,067±0,007 |
|
14 |
0,075±0,009 |
0,093±0,002 |
|
15 |
0,095±0,002 |
0,097±0,002 |
|
16 |
0,090±0,004 |
0,077±0,012 |
|
17 |
0,025±0,002 |
0,067±0,010 |
|
18 |
0,100±0,002 |
0,063±0,004 |
|
19 |
0,150±0,019 |
0,053±0,008 |
|
20 |
0,100±0,001 |
0,090±0,005 |
|
21 |
0,150±0,019 |
0,060±0,010 |
|
22 |
0,200±0,001 |
0,060±0,010 |
|
23 |
0,150±0,019 |
0,050±0,018 |
|
24 |
0,085±0,006 |
0,073±0,007 |
|
01 |
0,120±0,030 |
0,050±0,005 |
|
02 |
0,090±0,004 |
0,083±0,008 |
|
03 |
0,250±0,019 |
0,039±0,009 |
|
04 |
0,200±0,001 |
0,063±0,014 |
|
05 |
0,101±0,001 |
0,060±0,010 |
|
06 |
0,125±0,028 |
0,035±0,009 |
|
07 |
0.090±0.004 |
0,080±0,008 |
|
08 |
0,150±0,019 |
0,057±0,004 |
|
09 |
0,045±0,006 |
0,087±0,004 |
Таблица 5 -
|
Варианты опыта |
Мезор, нг/мл |
Амплитуда (пределы),нг/мл |
Акрофаза в часах и минутах(доверительный интервал) |
Перио д, ч |
|
Адреналин, норма |
0,047±0,003 |
0,013 (0,007÷0,020) |
21 ч 30 мин (19 ч 24 мин ÷01ч.06 мин) |
24* |
|
Адреналин, стресс |
0,036±0,004 |
0,016 (0,002÷0,31) |
16ч 42мин (13 ч 06 мин ÷23 ч.42 мин) |
25* |
|
0,017 (0,006 ÷ 0,031) |
01ч 36 мин (01 ч 06 мин ÷03 ч.24 мин), 14 ч 36 мин (14 ч 06 мин ÷16 ч.24 мин), |
13* |
||
|
Норадреналин, норма |
0,120±0,007 |
0,033 (0,020 ч- 0,042) |
02 ч 00 мин (00 ч 00 мин ÷ 03 ч 42мин) |
24* |
|
Норадреналин, стресс |
0,074±0,005 |
0,023 (0,010 ÷0,035 |
12 ч 36 мин (12 ч 00 мин ÷13 ч.30мин) |
24* |
|
0,019 (0,010 ÷0,039) |
11 ч 36 мин (8 ч 12 мин ÷12 ч 48.мин) 23 ч 36 мин (20 ч 12 мин ÷ 24 ч 48мин) |
12* |
||
|
Примечание: * р < 0,05 |
||||
Результаты Косинор – анализа суточной динамика катехоламинов в плазме крови крыс в норме и при стресс представлены в таблице 5.Из табл. 5 видно, что акрофаза 24 – часового ритма норадреналина также, как и адреналина, приходится на ночное время, в 02 часа ночи. Содержание катехоламинов в плазме крови, измеренное разные часы суток, показывает разнонаправленность их изменения при стрессе (см. табл. 3 и 4).
Так, содержание адреналина повышается в 13 часов, в 15- 16 часов и в 08 часов утра, содержание норадреналина повышается в 12 часов и в 14 часов, в остальное же время содержание этих гормонов в плазме крови крыс сохраняется на прежнем уровне либо значительно понижается. Мезор адреналина (см. табл. 5) в плазме крови крыс на вторые сутки гипокинезии снижается относительно нормы, также как и норадреналин. При стрессе наблюдается расщепление 24 – х часового периода на ультрадианные ритмы, т.е. на 12- ти и 13- ти часовые периоды.
Таким образом, нами впервые вычеслены значения хроноструктурных параметров (период, амплитуда, мезор, акрофаза) суточной динамики адреналина и норадреналина у крыс в норме и при стрессе и установлена отличительные особенности между ними.
Выводы:
- У здоровых кроликов существует хорошо выраженный синфазный одновершинный суточный (циркадианный ритм СЭП аурикулярных биоактивных точек кожи с акрофазой в полночь и минимумом в утренне – дневное время.
- Стресс, создаваемый экспериментальной острой ангиной, приводит к полифазности колебаний, увеличению мезора и амплетуды, возрастанию вариабельности, смещению значений на суточной шкале, потере однозначности показателей, расщеплению суточной динамики, СЭП на собственно 24- часовой и 08- часовой периоды.
- Концентрация гормонов надпочечников у интактных крыс имеет максимальное значение в ночное время, минимальное – в дневное время. Период суточного ритма составляет 24 часа. При гипокинезии (стрессе)у опытных крыс изменения суточных ритмов проходят несколько стадий. Суточный ритм удлиняется и составляет 24 часов,наблюдается снижение мезора и смещение акрофазы по шкале времени, расщепление суточных ритмовна ультрадианныес периодом 12-013 часов.
- Показано, что устойчивость организма животных к воздействию стрессовых факторов зависит от хроноструктурных параметров биоритмов. Стресс сопровождается резкими сдвигамисуточной (циркадианный) организации и хроноструктурных показателейэлектрических показателей биоактивных точек кожи и гормонов надпочечников у кроликов и крыс.
СПИСОК ЛИТЕРАТУРЫ
- Алякринский Б.С., Степанова С.И. По закону ритма. - М: Наука, 1985. – 174 с.
- Тулеуханов С.Т. Временная организация биологических систем.- Алматы: Қазақ университеті, 1999. – 157с.
- Тулеуханов С.Т., Ефимов М. Л. Хронобиологияи хрономедицина.- Алматы: Қазақ университеті, 1996.- 2003 с.
- Аблайханова Н.Т., Тулеуханов С.Т. Исследование хроноструктурных параметров временной организации электропроводности биоактивных точек кожи кроликов в норме и при адаптации к условиям гипоксии // Физиологические проблемы адаптации межрегион. конф. посв. 85-летию засл.деят. науки РФ, д.м.н., проф. Држевской И.А. – Ставрополь: 2008. - С. 190-192.
- Аблайханова Н.Т., Тулеуханов С.Т. Сравнительный анализ хроноструктурных параметров электрофизиологических показателей аурикулярных БАТ у животных в норме и после гипоксии // Фундаментальные исследования в биологии и медицине Сборник научных трудов. – Ставрополь: 2009. - С.-3-9.
- Aronson Doron. Impaired modulation of circadian rhythms in patients with diabetes mellitus: A risk faktor for cardiac thrombotic events // Chronobiol.Int. – 2001. – Vol.186, №1. – P. 109-121.
- Kawano Yuhei. Biorhythm and hypertension //Asion Med.J. - 2000. - Vol.436, №5. - C.207-213.
- Hermida Ramon C., Fernandes Jose R., Ayala Diana E., MojonArtemio,Al-onsoJgnacio, Smolensky Michael. Circadian rhythm of double (rate - pressure) product in healthy normotensive young subjects // Chonobiol. Jnt. – 2001.- №3(18). – Р. 475 - 489.
- Acuna-Goycolea C., Obrietan K., van den Pol A. N. 2010. Cannabinoids excite circadian clock neurons // J. Neurosis. – 2010. - №30. – Р. 10061-10066.
- Aguilar E. 2012. Kisspeptins and reproduction: physiological roles and regulatory mechanisms // Physiol. Rev. – 2012. - №92. – Р. 1235—1316.
- Тулеуханов С. Т., Ургалиев Ж. Ш., Бабашев А. Биологически активные точки наруженой ушный раковины кроликов и динамика их суточной активности // Генетические и биоэнергетические исследования организма. – Алма–Ата: КазГУ. 1982. – С.138-149.
- Тулеуханов С. Т. Некоторые вопросы исследования электрических свойстов «точки акупунктуры» кожи человека и животных в норме и патологии // Биологические науки. – Алма–Ата: КазГУ, 1974. - Вып. 7. – С. 120-124.
- Коган А. Б. Потенциалы кожи // Электрофизиология. - М.: 1969. - С. 141- 150.
- Лакомкин А. И., Мягков И. Ф. Электрофизиология. – М.: Высшая школа. 1997. - 232 с.
- Слынько П.П. Основы низкочастотной кондуктометрии в биологии. - М.:Наука,1972. – 132 с.
- Матлина Э. Ш. Флюорометрические методы определения адреналина и нораадреналина в крови и мочеlВКН // Адреналин и норадреналин. – М.: Наука, 1964. – С. 268- 276.
- ×åðíûøåâà M. Ï., Íîçäðà÷åâ À. Ä. 2006. Ãîðìîíàëüíûé ôàêòîð ïðîñòðàíñòâà è âðåìåíè âíóòðåííåé ñðåäû îðãàíèçìà. - ÑÏá.:Íàóêà. - 246 ñ.
- Ершов В.Ш., Сорокин А.А. Пакет прикладных программ Косинор – анализа и методические указания по его использованию // Информ.бюл. ГФАП СССР. – 1980. - №5.- С.38-45.
- Багриновский К. А., Багинская Н. В.,Баженова А. Ф., Колпаков М.Г., Романюха А. А., Маркель А. Л. Математический анализ циркадных систем организма на основаним процедуры «Косинор» // Кибернет. подходы к биологии. – Новосибирск: 1973. - С.196-209.
- Кривощеков С. Г., Матюхин В. А., Разумов А. Н., Труфакин В.А. Профилактика и прогнозирование десинхронозов: учеб. пособие. - М.: Новосибирск: СО РАМН, 2001. – 56 с.