В статье представлены современные сведения о внутриклеточных молекулярно-генетических механизмах, участвующих в возникновении и развитии остеоартрита (ОА). Освещена роль эпигенетической регуляции генов, Wntсигнального пути, а также взаимоcвязь процессов, происходящих в костной ткани, с остеоартритом.В статье суммированы открытия из этих областей и рассмотрено их возможное будущее применение в терапии.
Введение.
Остеоартрит (ОА) является наиболее распространенной болезнью суставов, вызывающий сильную боль, скованность, затруднение движения, отек, крепитацию, и приводящей к инвалидности. ОА характеризуется прогрессивным разрушением суставного хряща при совместном сужении межхрящевого пространства, вызывает формирование остеофитов, субхондральный склероз и синовит [1]. Коленной сустав является наиболее клинически значимым участком первичного очага ОА. Одним из современных методов оценки поражения сустава является рентгенологическая оценка, которая отражает тяжесть заболевания по степени дегенерации сустава. Существует так называемая оценочная шкала Келгрена-Лоуренса (англ. Kellgren-Lawrencegradingscale) [2]. Этиология и патогенез ОА остаются неясными, но их обычно связывают с несколькими физиологическими факторами, такими, например, как ожирение и старение [3]. Тем не менее, на данный момент точно известно, что биохимические факторы играют важную роль в возникновении и развитии ОА [4].
Исследования в фундаментальной биологии остеоартрита (ОА) продолжают расти вместе с увеличением работ касательно выяснении механизмов, лежащих в основе разрушения хрящевой ткани. В данном обзоре мы попытались обобщить все основные молекулярногенетические аспекты возникновения и развития остеоартрита.
Роль эпигенетической регуляции генов.
Эпигенетическая регуляции транскрипции генов позволяет клеткам изменять и настраивать экспрессию генов в ответ на внешние сигналы. В отличие от генетических модификаций, которые требуют очень длительного периода для возникновения, организмы могут использовать эпигенетические модификации для того, что бы более быстро реагировать на изменения окружающей среды. Отдельные клетки и ткани также могут использовать эпигенетические модификации с целью изменения экспрессии генов для быстрого реагирования на изменения в их микросреде. Многие эпигенетические модификации могут сохранятся в культивируемых клетках. Было замечено, что в течение некоторого времени хондроциты, удаленные из сустава, пораженного остеоартритом, и помещенные в культуральную среду, будут поддерживать фенотип ОА. Поддержание фенотипа наблюдается в течение недели или более, происходит экспрессия генов, наблюдающаяся при ОА, несмотря на то, что клетки находятся уже не в присутствии измененной среды сустава, пораженного ОА. Это явление может быть объяснено эпигенетическими изменениями в клетке, в том числе метилированием ДНК, метилированием гистонов, ацетилированием, микро (ми)РНК, которые регулируют экспрессию генов связанных с фенотипом ОА.
ММР-13 является критическим ферментом, который способствует разрушение хрящевого матрикса при ОА. Данный фермент находится под эпигенетической регуляцией. Увеличение экспрессии ММР-13 в хондроцитах человека при ОА связано с деметилированиемCpG сайтов в промоторе гена, кодирующего ММР-13 [5]. Было установлено, что при деметилировании данного промотора в положении -104 б.п. появляется участок связывающий белок, ответственный за связывание цАМФ (CREB). В параллельных исследованиях промотора гена ММР-13 деметилирование в положениях -110 б.п. и -299 б.п. показало связь с повышенным уровнем экспрессии гена ММР-13 [6]. В подобном исследовании обнаружили аналогичные сайты в промоторе гена IL-1b. Интересно, что связывание белка HIF-2а (который принимает участие в развитии ОА у мышей) с промотором гена ММР-13, но не связывающегося непосредственно с промотором гена IL-1b , усиливалось при деметилировании обоих промоторов.
Другой ген, связанный с катаболической деятельностью в хряще это окись азота индуцируемая синтаза (iNOS). Был обнаружен участок в энхансере NF-кВ, деметилирование которого ассоциированно с пониженной экспрессией гена данной синтазыв хондроцитах человека при ОА [7]. C анаболической стороны, метилирование промотора гена SOX-9 вызывало увеличение экспрессии синтазы в хондроцитах хряща человека при ОА по сравнению контролем [8]. Деметилирующий агент 5-азацитидин способен увеличить экспрессию гена SOX-9 демонстрируя, что метилирование является ингибирующим и это, как оказалось, опосредовано измененением связывания CCAAT- связывающий фактора / ядерного фактора-Y и фактора CREB. Метилирование и ацетилирование гистонов также может регулировать активность промотора гена SOX-9 . Увеличение уровня триметилирования в гистоновых белках H3K9 и H3K27 и снижение ацетилирования в H3K9, 15, 18, 23, и 27 показало взаимосвязь с регуляцией промотора гена SOX- 9.
Модификации гистонов может также участвует в регулировании катаболических медиаторов в хряще. Человеческие хондроциты, в которотых была индуцирована экспрессия генов Runx-2, ММР-3 и ADAMTS-5 посредством воздействия механического стресса, обрабатывали ингибиторами гистон деацетилазы (HDAC) MS-275 или A трихостатином (TSA) [9]. После данной обработки ингибиторами гистон деацетилазы наблюдалась пониженная экспрессия всех трех генов. Точно так же, IL-1- индуцированная экспрессия генов ММР-1, ММР-3, ММР-и 13 была уменьшена при обработке суставных хондроцитов человека ингибиторами TSA, MS-275 или вальпроевой кислотой [10]. В поддержку invivo роли ацетилирования гистонов в ОА, у модельных мышей с ОА, которых подвергали обработке ингибитором гистон ацетилазыTSA, наблюдалось сокращение тяжести повреждения хряща в дестабилизированном медиальном диске(DMM) [10]. Таким образом, на сегодняшний день все известные исследования указывают на сильную вовлеченность ряда катаболических и анаболических факторов в ОА хондроцитах, которые регулируется как посредством метилирования так и модификацией гистонов.
В дополнение к исследованиям в хряще, были исследованы различия в метилировании ДНК при ОА и метилировании в кости при остеопорозе, взятой из центральной части головки бедренной кости [11]. Из 241-го сайта в 228-ми генах, которые были дифференциально метилированны, для подавляющего большинства (217 сайтов) было установлено снижение метилирования данных сайтов в остеопоротичной кости. Когда функции этих генов были аннотированы, был установлен гомеобокс надсемейства транскрипционных факторов, который значительно отличается при ОА и при остеопорозе. Это является интересным наблюдением, учитывая важную роль этого семейства транскрипционных факторов в развитии костей и суставов.
Очень активной областью эпигенетических исследований являются исследования микро (ми)РНК, хотя какие именно (ми)РНК являются важными в биологии ОА до сих пор остается не вполне ясным. При микрочиповом анализе микроРНК, присутствующих в культуре нормальных и ОА хондроцитов человека, были обнаружены семь микроРНК, которые показали дифференциальную экспрессию в здоровых и ОА хондроцитах [12]. Только у одной miR483-5p наблюдалась увеличенная экспрессия в ОА клетках, в то время как у других (ми)РНК (miR-149, miR-582-3p, miR-1227, miR-634, miR576-5p и miR-641) наблюдался более высокий уровнень в нормальных хондроцитах. На данный момент не проводилось никаких исследований функций данных (ми)РНК, поэтому остается неясным какую роль играют данные (ми)РНК в патогенезе ОА. В другом исследовании, проведенном на человеческом хряще, было отмечено возраст-связанное увеличение уровня miR-199a-3p и miR193b [13]. В противоположность этому, хондроциты человека, подвергшиеся обработке IL-1b, имели сниженный уровень miR-199a, и это, как было показано, было связано с увеличением экспрессии циклооксигеназы-2 и последующей продукции PGE-2 [14]. Обработка иммортализованной линии хондроцитов (C28/I2) IL-1b приводила к повышенной экспрессии miRNA-140, которая, как было показано, ингибирует экспрессию MMP-13 [15]. Предыдущее исследование показало, что удаление miRNA-140 у мышей приводит к преждевременному проявлению ОА фенотипа в сочетании с увеличенной экспрессией Adamts5 [16].
Wnt сигнальный путь.
Wnt сигнальный путь играет ключевую роль в развитии сустава и последние работы поддерживают роль данного пути в ОА. Так как Wnt сигнальный путь участвует в процессах формирования как хряща так и кости, которые включают роль данного пути в регуляции гипертрофии хондроцитов в хрящевой пластинке, нарушение регуляции Wnt сигнального пути в тканях взрослого организма может способствовать гипертрофии хондроцитов, наблюдаемой при патологических изменениях в хрящевой и костной тканях при ОА. Склеростин служит в качестве ингибитора Wnt сигнального пути в кости и мутации в гене склеростина, приводящие к потере функциональности последнего, либо ингибирование склеростина посредством использования нейтрализующих антител, приводит к увеличению массы костной ткани. В дополнение к этому замечено, что склеростинэкспрессируется также в хрящевой ткани, но на одинаковом уровне как в здоровой ткани так и в ОА хондроцитах [17]. Тем не менее, ни генетическое удаление склеростина у мышей, ни ингибирование антителами у крыс не повлияло на естественное развитие ОА у мышей или на хирургически индуцированный ОА у крыс. Интересно, что у склеростиннокаутированных мышей 12-ти и 16-ти месячного возраста не наблюдались более тяжелые поражения хряща по сравнению с подобранными по возрасту контрольными мышами, несмотря на то, что у первых наблюдалось значительное увеличение массы субхондриальной кости. Эта находка выступает либо против прямого влияния повышенной массы субхондриальной кости на хрящевую ткань, либо предполагает, что потеря склеростина в хряще может иметь противоположный эффект на повышенние костной массы.
Белки Dkk 1-4 также являются антагонистами Wnt сигнального пути и, известно, что белки Dkk-1 и -2 присутствуют в суставном хряще, а Dkk-1 также обнаружен в синовиальной оболочке при ОА [18,19]. Замечено увеличение экспрессии Dkk-1 с одновременным уменьшением экспрессии Dkk-2 в ОА хрящевой ткани по сравнению с нормальной тканью [18]. В другом исследовании былы установлены более высокие уровни экспрессии DKK-1 и двух других антагонистов Wnt сигнального пути, белков Gremlin 1 и Frizzled-связанного белка в суставном хряще по сравнению с хрящевой пластинкой [20]. Было замечено, что данные антагонисты ингибируют гипертрофию хондроцитовinvitro, что говорит в пользу того, что более высокие уровни данных белков в суставном хряще могут служить для ингибирования гипертрофии. Индукция гирерэкспрессии Dkk-1 посредством инъекции вектора на основе аденовируса в сустав мышей привела к снижению тяжести ОА в DMM модельных мышах [18]. Тем не менее, был замечен противоположный эффект Dkk-1 на ОА у модельных крыс ACLT, где интраперитонеально производилась инъекция антисмысловыхолигонуклеотидов к матричной РНК Dkk-1, достаточных для уменьшения количества DKK-1- положительных клеток в синовиальной оболочке, что, в свою очередь, привело к снижению степени тяжести ОА [19]. Было установлено, что Dkk-1 ингибирует экспрессию ММР- 13 и ADAMTS-4 в хондроцитах в ответ на активаторWnt сигнального пути Wnt-3a [18], что, в свою очередь, способствует производству про-ангиогенных факторов и протеиназ (ADAMTS-5 и ММР-3) синовиальными фибробластами [19]. Если эти данные о противоположных эффектах в хряще и синовиальной оболочке подтвердятся, то это будет говорить о том, что использование Dkk-1 как мишень для лечения ОА будет невозможно при поражении этих двух тканей.
Также усложняет понимание роли Wnt сигнального пути в развитии ОА, исследование, при котором была обнаружена повышенная экспрессия Wnt-1-индуцируемого сигнального белка 3/CCN6 в ОА хрящевой ткани по сравнению с нормальной хрящевой тканью, и в invitroэкспериментах было обнаружено, что данный сигнальный белок подавляет экспрессию ADAMTS-5 и, в то же время усиливает экспрессию MMP-10 [21]. Ингибирование активации Wnt /b- катенинового сигнального пути впервичныххондроцитах человека, происходящее посредством стимуляции IL-1b белков ММР-1, -3, -13, и, возможно, этот каскад происходит путем ингибирования транскрипционного фактора NFkB [22]. Тем не менее, другие invitroисследования указывают, что белки семейства Hedgehog стимулируют накопление протеогликаналубрицина, в то же время, антагонисты Wnt / B-катенинового сигнального пути также стимулируют накопление лубрицина [23]. Эти исследования показывают нам, что для создания оптимальных и точных терапевтических мишеней, необходимо более полное и глубокое понимание регуляции Wnt сигнального пути в тканях сустава.
Костная ткань.
Роль кости в биологии ОА является активной областью исследований. Хотя изменения в суставном хряще при ОА у некоторых людей и в животных моделях (см. выше исследования на DMM модельных мышах) можно заметить даже при отсутствии значительного синовита, вовлечение костной ткани вблизи пораженного сустава в развитие ОА возможно является универсальным по крайней мере на некоторых стадиях развития ОА. Тем не менее, различные исследования показывают как сильную корреляцию между изменениями, происходящими в человеческой хрящевой ткани при ОА и животными моделями, так и почти полное отсутствие последней между ними. В образцах человеческой остеохондральной ткани, взятых из большой берцовой кости рядом с поврежденным хрящем, при помощи микротомографии (micro-CT) были обнаружены изменения в степени минерализации и объемах костной ткани, и последние параметры находились в жесткой корреляции со степенью поражения хрящевой ткани [24]. Этой же группой исследователей было опубликовано второе исследование роли вклада чрезмерной нагрузки и эндохондрального окостенения в субхондральные изменения в костной ткани при ОА, что привело их к выводу, что оба данных процесса ответственны за изменения в костно-хрящевой ткани [25].
В исследованиях, проведенных в различных мышиных ОА моделях, отмечены как сильная так и слабая корреляция между изменениями в хрящевой и костной тканях [26]. Гиперэкспрессия продукта гена EphB4 в костной ткани приводила к уменьшению уровня повреждения хряща в DMM модели, что коррелирует со снижением субхондрального склероза и объема кости, а также с количеством фосфатаза-положительных остеокластов [27]. Удивительно, что в отличие от этих исследований, сравнение с контрольной группой дикого типа мышей с Col6a1-/- мышами показало значительное уменьшение потерь хрящевой ткани коленного сустава в возрасте 15 месяцев, несмотря на большие остеофиты, которые появились в возрасте уже 2 месяцев [28]. У склеростин нокаутированных мышей не наблюдалось возрастного проявления ОА на 12 или 16 месяце, несмотря на значительное увеличение массы субхондральной кости [17]. Различия в формировании остеофитов и в повреждении хрящевой ткани были также отмечены в исследованиях, изучающих влияние ингибирования катепсина К на развитие ОА. Катепсина К – протеаза, продуцируемая остеокластами и вовлеченная в разрушение костной ткани. Также было установлено, что катепсин К производится синовиальными фибробластами и хондроцитами. В недавних исследованиях было обнаружено, что катепсин К может способствовать расщеплению коллагена типа II, ассоциированного со старением и с OA [29]. У модельных кроликов, подвергшихся обработке химическим ингибитором катепсина К (L-235), наблюдалось уменьшение повреждение хряща как и у катепсин К нокаутированных мышей по шкале H. Mankin [30]. В обеих моделях ингибирование катепсина К способствовало защите субхондральной кости. Тем не менее, значительное снижение остеофитов наблюдалось только в модельных кроликах, где химический ингибитор лишь частично защищал от повреждения хряща, в то время как в катепсинК-/- мышах не наблюдалось сокращения остеофитов, наряду с очень значительным снижением повреждений по шкале H. Mankin.
Заключение.
Необходимо изучение различных тканей, вовлеченных в процесс возникновения и развития ОА, и являющихся поставщиками потенциальных медиаторов, которые вносят вклад в развитие ОА (рисунок 1). Конечной целью исследований биологии ОА является открытие новых терапевтических мишеней на основе более детального понимания механизмов болезни. Хотя эпигенетическая регуляция транскрипции генов и Wnt сигнальный путь вносят свой вклад в процесс ОА, вся сложность взаимодействующих систем приводит к трудностям в рассмотрении деятельности одного аспекта, которая может быть полезной при изучении ОА, не затрагивая другой аспект, который также влияет на течении ОА. Поэтому переплетение регулятивных путей, их взаимосвязь делает эти области трудными для подбора новых терапевтических решений. Но это не значит, что данные аспекты не являются важными для дальнейшего исследования, это лишь указывает нам на то, что применение вновь полученных знаний в данной области требует более точного понимания вопроса и, таким образом, разработка и внедрение новых точных терапевтических мишеней в клинику потребует более длительного периода по сравнению с другими областями.
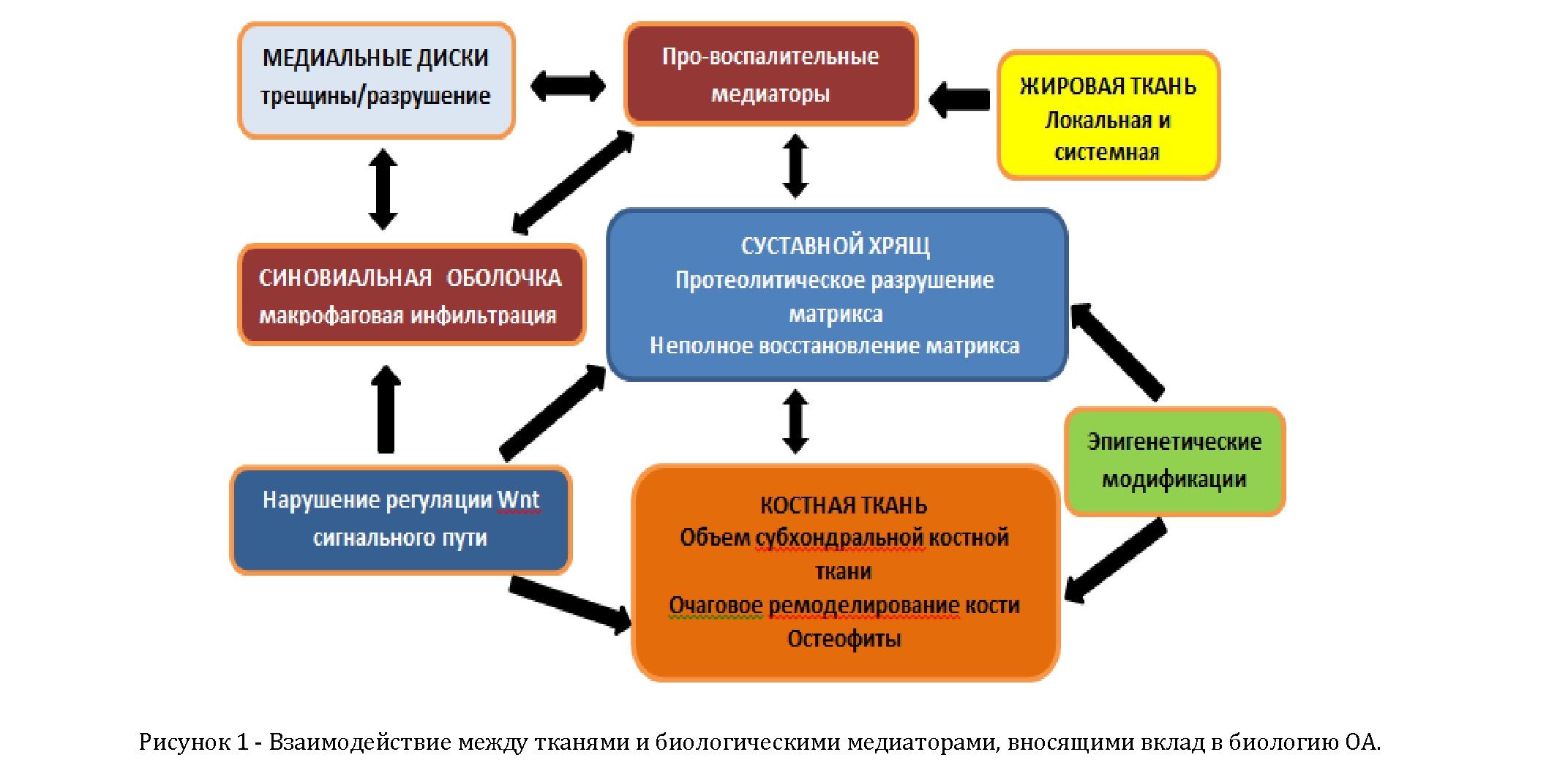
Знания о роли воспаления в ОА быстро расширяются и есть свидетельства о том, что про-воспалительные медиаторы продуцируются всеми суставными тканями, пораженными ОА. Ограничением в данной области исследований является то, что нет четко определенного отдельного целевого агента, который возможно было бы использовать в качестве мишени, в отличие, например, от ревматоидного артрита
(РА), где таргетинг TNFa приводит к значительному облегчению симптомов заболевания у многих (но не у всех) пациентов. При ОА это может быть набором неких провоспалительных медиаторов, которые необходимо будет ингибировать. Это может быть вполне возможно, как отмечено в работе по исследованию хемокинового рецептора CCR2, где показано, что провоспалительные медиаторы, ответственные за возникновение боли отличаются от медиаторов, ответственных за разрушение ткани сустава.
Наконец, в данном обзоре были рассмотрены также вопросы взаимосвязи костной ткани с остеоартритом. Тем не менее, существующие данные не указывают на последовательную связь между изменениями в костной ткани и разрушением соседствующей хрящевой ткани, в частности в отношении формирования остеофитов. Это говорит о том, что в данном случае мы можем наблюдать аналогичную ситуацию как при воспалении в ОА, то есть возможно существование таких вариантов возникновения ОА, где костная ткань является иницирующим фактором в процессе становления и развития болезни. И ясно, что в данном случае, остеофиты не являются лучшим маркером. Также важно упомянуть о важности исследований локальных процессов, происходящих при ремоделировании кости, которые стало возможным изучать с помощью методов визуализации, таких, например, как МРТ, и, которые могут быть важны при возникновении боли и/или развитии заболевания. Очевидно, что знания в понимании процессов, происходящих при остеоартрите,стремительно растут, но лучшее понимание биологии данного заболевания, которое поможет исследователям осуществить переход на принципиально новые виды терапии, требует разделения остеоартрита на различные типы, которые имеют, в свою очередь, различную этиологию.
СПИСОК ЛИТЕРАТУРЫ
- Ralston, SH (2002) Genetic control of susceptibility to osteoporosis.// J. ClinEndocrinolMetab 87: pp. 2460-2466.
- Hill, TP, Später, D, Taketo, MM, Birchmeier, W, Hartmann, C (2005) Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. DevCell 8: pp. 727-738.
- Baron, R, Rawadi, G (2007) Targeting the Wnt/ß-catenin pathway to regulate bone formation in the adult skeleton. //Endocrinology 148: pp. 2635-2643.
- Glass, DA, Bialek, P, Ahn, JD, Starbuck, M, Patel, MS, Clevers, H, Taketo, MM, Long, F, McMahon, AP, Lang, RA, Karsenty, G (2005) Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev Cell 8: pp. 751-764.
- Bui C, Barter MJ, Scott JL, Xu Y, Galler M, Reynard LN, et al. cAMP response element-binding (CREB) recruitment following a specific CpGdemethylation leads to the elevated expression of the matrix metalloproteinase 13 in human articular chondrocytes and osteoarthritis. FASEB J 2012;26:3000-11.
- Hashimoto K, Otero M, Imagawa K, de Andres MC, Coico JM, Roach HI, et al. Regulated transcription of human matrix metalloproteinase 13 (MMP13) and interleukin-1beta (IL1B) genes in chondrocytes depends on methylation of specific proximal promoter CpG Sites. J BiolChem2013;288:1006-72.
- Andres MC, Imagawa K, Hashimoto K, Gonzalez A, Roach HI, Goldring MB, et al. Loss of methylation in CpG sites in the NF kappaB enhancer elements of inducible nitric oxide synthase is responsible for gene induction in human articular chondrocytes. Arthritis Rheum 2013;65:732-42.
- Kim KI, Park YS, Im GI. Changes in the epigenetic status of the SOX-9 promoter in human osteoarthritic cartilage. //J Bone Miner Res 2013;28:1050-60.
- Saito T, Nishida K, Furumatsu T, Yoshida A, Ozawa M, Ozaki T. Histone deacetylase inhibitors suppress mechanical stress-induced expression of RUNX-2 and ADAMTS-5 through the inhibition of the MAPK signaling pathway in cultured human chondrocytes. Osteoarthritis Cartilage 2013;21:165-74.
- Culley KL, Hui W, Barter MJ, Davidson RK, Swingler TE, Destrument AP, et al. Class I HDAC inhibition modulates metalloproteinase expression and blocks cytokine-induced cartilage degradation. Arthritis Rheum 2013.
- Delgado-Calle J, Fernandez AF, Sainz J, Zarrabeitia MT, Sanudo C, Garcia-Renedo R, et al. Genome-wide profiling of bone reveals differentially methylated regions in osteoporosis and osteoarthritis. Arthritis Rheum 2013;65:197-205.
- Diaz-Prado S, Cicione C,Muinos-Lopez E, Hermida-Gomez T, Oreiro N, Fernandez-Lopez C, et al. Characterization of micro-RNA expression profiles in normal and osteoarthritic human chondrocytes. BMC MusculoskeletDisord 2012;13:144.
- Ukai T, Sato M, Akutsu H, Umezawa A, Mochida J. MicroRNA-199a-3p, microRNA-193b, and microRNA-320c are correlated to aging and regulate human cartilage metabolism. J OrthopRes 2012;30:1915-22.
- AkhtarN, Haqqi TM. MicroRNA-199a* regulates the expression of cyclooxygenase-2 in human chondrocytes. Ann Rheum Dis 2012;71:1073-80.
- Liang ZJ, Zhuang H, Wang GX, Li Z, Zhang HT, Yu TQ, et al.MiRNA-140 is a negative feedback regulator of MMP-13 in IL-1beta-stimulated human articular chondrocyte C28/I2 cells. Inflamm Res 2012;61:503-9.
- Miyaki S, Sato T, Inoue A, Otsuki S, Ito Y, Yokoyama S, et al. MicroRNA-140 plays dual roles in both cartilage development and homeostasis. Genes Dev 2010;24:1173-85.
- Roudier M, Li X, Niu QT, Pacheco E, Pretorius J, Graham K, et al. Sclerostin is expressed in articular cartilage but loss or inhibition does not affect cartilage remodeling during aging or following mechanical injury. Arthritis Rheum 2012; 65:721-31.
- Oh H, Chun CH, Chun JS. Dkk-1 expression in chondrocytes inhibits experimental osteoarthritic cartilage destruction in mice. Arthritis Rheum 2012;64:2568-78.
- Weng LH, Ko JY, Wang CJ, Sun YC, Wang FS. Dkk-1 promotes angiogenic responses and cartilage matrix proteinase secretion in synovial fibroblasts from osteoarthritic joints. Arthritis Rheum 2012; 64:3267-77.
- Leijten JC, Emons J, Sticht C, van Gool S, Decker E, Uitterlinden A, et al. Gremlin 1, frizzled-related protein, and Dkk-1 are key regulators of human articular cartilage homeostasis. Arthritis Rheum 2012;64:3302-12.
- Baker N, Sharpe P, Culley K, Otero M, Bevan D, Newham P, et al. Dual regulation of metalloproteinase expression in chondrocytes by Wnt-1-inducible signaling pathway protein 3/CCN6. Arthritis Rheum 2012;64:2289-99.
- Ma B, van Blitterswijk CA, Karperien M. A Wnt/beta-catenin negative feedback loop inhibits interleukin-1-induced matrix metalloproteinase expression in human articular chondrocytes. Arthritis Rheum 2012;64:2589-600.
- Wei F, Zhou J, Wei X, Zhang J, Fleming BC, Terek R, et al. Activation of Indian hedgehog promotes chondrocyte hypertrophy and upregulation of MMP-13 in human osteoarthritic cartilage. Osteoarthritis Cartilage 2012;20:755-63.
- Cox LG, van Donkelaar CC, van Rietbergen B, Emans PJ, Ito K. Decreased bone tissue mineralization can partly explain subchondral sclerosis observed in osteoarthritis. Bone 2012; 50:1152-61.
- Cox LG, van Donkelaar CC, van Rietbergen B, Emans PJ, Ito K. Alterations to the subchondral bone architecture during osteoarthritis: bone adaptation vs endochondral bone formation. Osteoarthritis Cartilage 2013;21:331-8.
- Hashimoto S, Rai MF, Janiszak KL, Cheverud JM, Sandell LJ.Cartilage and bone changes during development of posttraumatic osteoarthritis in selected LGXSM recombinant inbred mice. Osteoarthritis Cartilage 2012;20:562-71.
- Valverde-Franco G, Pelletier JP, Fahmi H, Hum D, Matsuo K, Lussier B, et al. In vivo bone-specific EphB4 overexpression in mice protects both subchondral bone and cartilage during osteoarthritis. Arthritis Rheum 2012;64:3614-25.
- Christensen SE, Coles JM, Zelenski NA, Furman BD, Leddy HA, Zauscher S, et al. Altered trabecular bone structure and delayed cartilage degeneration in the knees of collagen VI null mice. PLoS One 2012; 7:e33397.
- Dejica VM, Mort JS, Laverty S, Antoniou J, Zukor DJ, Tanzer M, et al. Increased type II collagen cleavage by cathepsin K and collagenase activities with aging and osteoarthritis in human articular cartilage. Arthritis Res Ther 2012;14:R113.
- Hayami T, Zhuo Y, Wesolowski GA, Pickarski M. Duong le T. Inhibition of cathepsin K reduces cartilage degeneration in the anterior cruciate ligament transection rabbit and murine models of osteoarthritis. Bone 2012;50: 1250-9.