In this article the results of the primary introduction of rare species of the genus Dactylorhiza Nevski in the conditions of the mountain forest zone of Eastern Kazakhstan are presents. As a result of field expedition work of 2017-2018, on the ridges of Ivanov and Kalba in the Kazakhstan section of the Altai mountain system, the following species of Dactylorhiza were added to the Altai Botanical Garden collection: D. fuchsii (Druce) Soo, D. longifolia (L. Neum.) Aver., D. incarnata (L.) Soo, D. maculata (L.) Soo, and D. umbrosa (Kar. & Kir.) Nevski. Survival rate was assessed under the conditions of introduction. The description of the external structure of generative individuals added to the culture was recorded by morphological characteristics and morphometric parameters: the shape and size of the inflorescences and leaves and the number and structure of flowers. The morphological characteristics and morphometric indicators of individuals of species added to the culture are the starting indicators when studying the introduction potential of the studied species of the genus Dactylorhiza. The collection of species of the genus Dactylorhiza will serve as the basis for the study of genetic diversity, molecular genetic analysis of species and intraspecific variability, which in turn will help to accurately distinguish between overlapping species.
Dactylorhiza Nevski is one of the widespread and numerous genera of the Orchid family of the temperate zone. The genus includes 50-70 species found in countries with a temperate and boreal climate, mainly in non-tropical Eurasia [1]. It is not yet fully researched and still not fully developed as a group [2]. Representatives of the genus are distinguished by high phenotypic variability, often hybridizing with each other, which makes it impossible to clearly distinguish the species even within the same coenopopulation or closely located communities [3, 4].
Most species of the genus, according to the gradation of R.V. Camelin [5], are classified as vulnerable and endangered, or, infrequently, rare.
In the «Flora of Kazakhstan» [6], the genus Dactylorhiza has 8 independent species, of which the EuroSiberian species D. fuchsii (Druce) Soo is included in the «Red Book of the Republic of Kazakhstan» [7], meaning that it's considered endangered.
The introduction of rare plant species is the main and most reliable way to preserve biodiversity. According to the works of S.A. Mamaev [2] and P.G. Efimov [8], species of the genus Dactylorhiza tolerate transplantation quite well and, when creating similar natural environmental conditions, grow equally well. As a result of field botanical surveys in the mountainous system of Kazakhstan Altai, initiated in 2017, and continued in 2018 as part of the grant project AP05133868 «Study of the distribution and current state of populations of Orchid species of the Kazakhstan Altai and their introduction in the Altai Botanical Garden» live plant samples of the genus Dactylorhiza from the family Orchidaceae (Lindl.) were displayed in the Botanical Garden's exposition of the natural flora of the Altai mountains (Ridder).
The purpose of this study is the morphometric characterization of samples of species of the genus Dactylorhiza from the Kazakh Altai, selected for primary introduction to the Altai Botanical Garden. The information reflected in this article can serve as a basis for comparison with individuals in introduction populations.
Materials and methods
The description of the external structure of the plant was carried out according to the methods of A.L. Fedorov and Z.T. Artyushenko [9-12]. The color palette of the inflorescences is determined on the basis of A.S. Bondartsev's color scale. [13] and the color scale of the English Royal Society of Florists [14]. Processing of statistical results was carried out according to the methods of G.N. Zaitsev [15] and A. Dospekhov [16]; Latin names were assigned to the plants in accordance with the nomenclature report S.К. Cherepanov [17].
The status of the species, depending on the endangered state and the category of rarity, is determined in accordance with the classification proposed by R.V. Kamelin and G.G. Sokolova in the Red Book of the Altai Region [5].
The Altai Botanical Garden is located on the south-eastern outskirts of the town of Ridder in the mountain-forest zone of the south-western periphery of the Western Altai. The limiting factors for the introduction of plants in a botanical garden are sudden changes in temperature and humidity throughout the year, season and day, and a short growing season. The climate of the area of introduction is characterized as sharply continental with a long cold winter and hot summer and is located in to a moderately cool, moderately humid agroclimatic zone. October is the autumn-winter transition period in the area of introduction. In October, the temperature in the pre-dawn hours is -3 °C to -5 °C; over the course of the day, air temperature drops from +16 °C to -10 °C. The winter period begins in first ten days of November, when the average daily temperature consistently drops by 5 °C. Soil freezing varies from 40 to 90 cm. According to the Ridder weather station, the frost-free period is 95-125 days. A sharp increase in temperature begins in the last week of May and reaches a maximum in July or occasionally in June. The average annual precipitation ranges from 432 mm to 937 mm with the greatest precipitation in summer. Soils are black dusty loams (humus content — 6.4 %) [18].
Research results and discussion
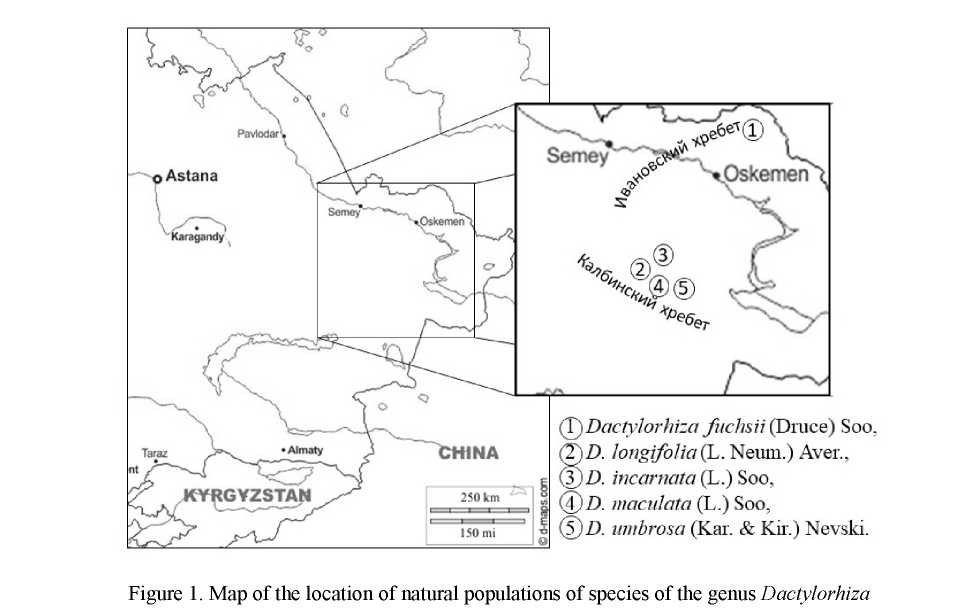
For the initial introduction of natural populations (Fig. 1) the collection consisted of the following species from the genus Dactylorhiza: D. fuchsii (Druce) Soo, D. longifolia (L. Neum.) Aver., D. incarnata (L.) Soo, D. maculata (L.) Soo, and D. umbrosa (Kar. & Kir.) Nevski.
Dactylorhiza fuchsii (Druce) Soo (Fig. 2) is a vulnerable meadow-forest species, with a shrinking habitat. D. fuchsii is reminiscent of relicts from the Paleogene of the taiga [19]. The species is classified as rarity category 2b or v (b), meaning that the species is vulnerable and has a limited range. The species bears a Protected status on the state level. D. fuchsii in the Kazakh Altais grows in small populations along the bed of mountain streams, under the canopy of mixed forests. D. fucksii plants occupy open areas that are not occupied by grass and that are richly moist, but without noticeable stagnation of water. Soils, as a rule, are abundantly humified, well-drained and alkaline. D. fucksii forms small groups of 3-5 individuals, often found in single specimens among rocky protrusions on moss cushions in communities with meso-hygrophilous spe-
Серия «Биология. Медицина. География». № 2(94)/2019
37
cies: Vacinium myrtillus L., Equisetum sylvaticum L., Geranium albiflorum Ledeb., Geum rivale L., Pyrola rotundifolia L., Carex macroura Meinsh., G. boreale L., Solidago virgaurea L., Draba sibirica (Pall.) Thell., Thalictrum flanum L., and Veronica longifolia L.
In the Altai Botanical Garden, the species is displayed on 25 grass turfs of various age states from the north-western foot of the Ivanov Ridge. Coordinates of location: 50º18' N, 83º35' E, 725 m above sea level. On the experimental plot, 3 form patterns were planted, differing in morphological variations.
Form pattern 1. Individuals with pure white flowers (very rare), undersized, leaves mostly without spots and specks.
Form pattern 2. Species with light pink flowers (widespread), medium-grown, leaves with dull pale purple specks of rounded shape, sometimes with dots.
Form pattern 3. Individuals with purple flowers, tall (most common), leaves with pale purple or deep purple spots, concentrated at the base of the leaf.
The shape of the leaves in all form patterns of D. fuchsii is obovate and slightly pointed at the end. In one generative adult individual, there are 1-4 (x = 2.43, hereafter is the average index) fully formed basal leaves, always blunt and rounded at the end. Less commonly, generative models show abundant formation of a large number of basal leaves of the whorled type. These organisms showed stems with 2-4 (x = 3.37) upward-directed lanceolate or narrow obovate leaves. Juveniles display narrow-lanceolate leaves. Under the inflorescence 1-3 (x = 2) narrowly linear underdeveloped bracts are located.
Visually established characteristic dependence: the darker the spots on the leaves, the richer the color of the inflorescence. In individuals involved in the introduction, the inflorescences, as a rule, are small, but there are also large ones from conoid to oblong-cylindrical in shape, depending on the age condition. The protruding lower lip of the flower is always clearly tripartite (trident), with a lighter colored middle section that is obtuse or wedge-shaped at the end and has longitudinal, maroon specks. In young, first flowering individuals, the middle lobe of the lower lip usually does not protrude, is very narrow and has a sphenoid shape. A well-developed spur always has a cylindrical shape; its length reaches 5-7 mm. Stipules are usually shorter than the flower.
The height of individuals in the sample with white flowers does not exceed 12-22 cm. The inflorescences are shortened, 3 cm long and 2 cm wide, and have a conical shape. An average of 26 flowers formed per individual. Bracts are 5-7 mm long and protrude a bit from the flowers. Basal leaves reach 7-8 cm in length; the width at the base of the leaf is 1-1.5 cm. In contrast to the basal leaves, the stem leaves are smaller; their length reaches 6 cm and width 5-6 mm. Bracts under the inflorescence have a lanceolate form, and are not more than 2 cm in length and 3 mm in width.
The white color variation of the inflorescence is a rather rare phenomenon, presumably associated with the absence of a dark coloring pigment.
Individuals with a light pink color of inflorescences are widely distributed within the species, characterized by relatively high competitive ability, unlike other species of this genus. As a rule, this variation is found in low-grass communities along the banks of streams, on moss beds among the stones.
The survival rate of form pattern in the year of planting on abundantly humified, loose soils with regular irrigation during the month was 100 %. In nature, with dense herbage, the height of individuals varies from 16 to 23 (x = 20.25) cm. At the base, the stems do not exceed 3-4 mm in diameter. The inflorescences are shortened, cylindrical with a length of 2-3.5 cm and a width of 1.5-2 cm. On average, one inflorescence consists of 30 flowers. All lower flowers are protected by narrow bracts, which vary in color from green to slightly brownish. Bracts distinctly protrude from the inflorescence. The lower leaves reach maximum sizes:

7—13 (х = 10) mm long and 3—4 (х = 3.5) mm wide. It should be noted that in individual mature plants of form pattern 2, individuals were found with numerous obovate radical leaves forming rosette whorls. The length of the basal leaves varies from 7.5—11 (х = 8.87) cm and width 1.2—1.5 (х = 1.42 ± 0.2, Cv = 10 %, hereinafter Cv — the coefficient of variation). Stem leaves are narrow and lanceolate in form, pointed at the end, with dim specks, from 6 to 12 (х = 8.25) cm long and 4—5 mm wide. Bracts are much smaller than other leaves: 1.1—2.5 (х = 1.8 cm) in length, always pointed, narrow-lanceolate, no more than 1—2 mm wide, green in color, and sometimes have a thin purple border around the edge.
Individuals with a violet inflorescence are the tallest — 30—47 (х =36) cm. They are most often located in tall grass meadow communities or on the marshy forest edges. Stems are solid, without cavities, and measure 0.7—0.9 (х = 0.83) cm in diameter at the base. This form pattern is characterized by long cylindrical inflorescences, from 5.5 to 9 (х = 6.8) cm long and 2—3.5 (х = 2.66) cm wide. The flowers are well formed, without anomalies. On average, one inflorescence consists of 46 flowers. The protruding bracts effectively protect the flowers from mechanical damage. Lower bracts are lanceolate or narrow-wedge-shaped in form, green in color and reach 6—15 (х = 10.25) mm in length. At the base of the stems are 3—4 obovate basal leaves, reaching a length of 9—16 (х = 11.5) cm, and a width of 1.5—3 (х = 2) cm. The stalks are seated with spotty lanceolate or narrow lanceolate leaves, 6—10 (х = 9) cm long and 1—1.5 (х = 1.3) cm wide. Under the inflorescence there are two bracts, with a brownish border around the edge, which is occasional absent. The length of these leaves does not exceed 2.5—3.5 (х = 2.8) cm, nor the width 2—4 (х = 3) mm.
Dactylorhiza incarnata (L.) Soo (Fig. 3) is a meadow-marsh rare species found in the Euro-Asian paleoarctic range [1, 20]. Its Rarity category: 3b or R (b) — a rare species with a wide range, often growing in specific habitats. Protection status: local protection. According to research by a number of scientists [21, 22], it has a narrow ecological amplitude. Research of V.G. Sobko [23] on rhizorestorative reproduction proves the ability of the species to vegetative propagate. A.A. Alekhin [24] considers D. incarnata a promising species for introduction.
On the Kalba ridge of the Kazakhstan Altai, the species is widespread, but not abundant. As a rule, its growth is confined to floodplain meadows, forest edges with partial shading, along the valleys of rivers and streams, and in tall grasses. The species does not tolerate complete shading as well as strong insolation. In plant communities, this species has an average competitive ability, but is often supplanted by turf grasses and sedges, especially when the ecology changes.
The main populations of D. incarnata are found in the grassy meadows in the foothills of the Kalba ridge. Unfortunately, this species is experiencing a strong anthropogenic impact due to overgrazing of livestock and summer mowing before the fruiting phase.
In surveyed regions of foothills in Kalba ridge, in additional to the main seed propagation, the vegetative method was achieved by planting auxiliary digitate tubers. In the populations of the species, the leftsided age spectrum prevails due to the predominance of immature and virginal individuals.
For the initial introduction into the Altai botanical garden D. incarnata was introduced by living plants from the mountain-meadow communities of the Kalba ridge. Coordinates of location: 49 ° 35´ N, 82 ° 31´ E, 634 m. For planting in culture, 20 different-age generative individuals were removed from nature, which were planted in the exposition of natural flora in both completely open and partially shaded areas with well humified, mildly acidic black soil substrate.
At abundant soil moisture without stagnation of water on the surface, a high survival rate of individuals was noted.
Серия «Биология. Медицина. География». № 2(94)/2019
39

By individual morphometric indices the specimens of D. incarnata, planted in the botanical gardens was 25–41 (x = 36 cm) high, with a thin, brittle, partially hollow stem 7–9 mm thick at the base. Inflorescences are cylindrical in shape, well-formed, and without anomalies in structure. Color variations range from dark pink to deep dark crimson. The size of the inflorescences reaches 6.5–7.5 (x = 7) cm in length and 2.5–3 (x = 2.8) cm in width. With strong insolation, the inflorescence becomes brownish. 36 to 42 (x = 37) flowers form per inflorescence. The lower floral lip is almost solid, diamond-shaped, with a narrow middle part slightly protruding forward by 1.5–2 mm. The length of the lip is almost equal to its width. The middle lobe of the lower lip is lighter than the lateral periphery, surrounded by a clear dark border, seated with numerous specks, and is round or wedge-shaped at the end. The oblong spur is to 7–9 mm long, conical in shape, and always lighter than the lip. Each flower is protected by a bract of light violet color, which is always longer than a flower. The bracts of the lower flowers are much larger than the upper ones: up to 17–20 (x= 20) mm in length and 4–6 (x= 5) mm in width.
The stem is covered with light green-keeled leaves, which are without spots, lanceolate or narrow- lanceolate, and have a maximum width below the middle. At the base of the stem is one basal leaf, 11 to 15 (x = 13.1) cm in length and 2–3 (x = 2.7) cm in width. There are 2–3 stem leaves as a rule, which are directed upward, lanceolate, and always pointed. The length of the stem leaves varies from 6 to 10 (x = 8.5) cm, width 1–1.5 (x= 1.33) cm. 1–3 (x = 2) shortened narrow lanceolate leaves are located under the inflorescence. Bracts do not exceed 3–4 (x = 3.5) cm in length or 5–6 (x = 5.7) mm in width, and as a rule, have a purple border around the edge.
Dactylorhiza longifolia (L. Neum.) Aver. (Fig. 4) — Eurasian, polymorphic species. Rare category: 2c or v (c) — a vulnerable view with a wide range. Protection status: local protection. D. longifolia has a hybrid origin — D. fuchsii (Druce) Soo × D. incarnata (L.) Soo [25]. The plant forms with a rather interesting rich color of the inflorescence and erect leaves with clear black spots. The species was found in the darkened forest edges, on abundantly wetted, marshy meadows in a community with grasses, and less often with sedge, though it can also be met under conditions of full light [26].
In Kazakh Altai mountain region D. longifolia grows sparsely, and populations are small. The collection was introduced as living plants from the Kalba ridge, from the southern part of the Sibin depression. The population grows on the bank of a small stream under a canopy of sparse birch with an admixture of willow; the soils are well humified due to rotten bedding.
Coordinates of location: 49º 40´ N, 82º 14´ E, 624 m above sea level.
For the primary introduction we collected 3 generative individuals of different ages. Since the species is very sensitive to environmental changes, especially to moisture, for successful adaptation of experimental plants to a botanical garden, a site was selected according to conditions as close as possible to natural ones, which ensured 100 % survival rate after proper care.
The plants are on average 30 cm tall, with relatively strong stems. Inflorescences have a rich dark crimson color with a cylindrical, less conoid form up to 6 cm long and up to 3 cm wide. On average, there are per inflorescence 35 flowers. Flowers from the collection are all properly formed. Anomalies were not found. The lower lip is indistinctly tripartite, to a solid diamond shape, and is always lighter at the base. The length of the lower lip is almost equal to its width. The middle lobe of the lower lip is rather narrow, has a wedgeshaped or infrequently rounded form, with longitudinal dark linear specks. Spur under the lower lip is elongated, 6–8 mm long, cylindrical, slightly flattened laterally. Wedge-shaped bracts that protrude from the inflorescence by 3–4 mm protect flowers from adverse factors.

The color of leaves varies from dark green to bluish-green, with numerous spots. The spotting on the leaves is always clear, without blurring, and well distinguishable. Spots are roundish, brown, distributed evenly over the entire area of the leaf. The shape of the leaves varies greatly from broadly lanceolate basal to lanceolate stem. A notable feature of all leaves is the sharpness at the end. Stems are dark, brownish, sometimes green, hollow, with defects, and with a base 10 mm thick, having low foliage. At the base of the stem is one broadly lanceolate leaf, 9-15 (x = 13) cm long and 3-5 (x = 4) cm wide. 2 to 4 lanceolate to narrow- lanceolate stem leaves per generative shoot are formed, always directed upward, 6-9 cm long, and 1.5 cm wide. Two lower bracts are present per plant, are rather narrow, are always longer than the bracts in the flower, and may have a brownish tint. The size of the upper stem leaves varies from 3.5 to 4 cm in length and from 4 to 6 cm in width.
Dactylorhiza maculata (L.) Soo (Fig. 5) is a rare species with a Euro-Siberian habitat. Rare category: 2c or v (c) — a vulnerable outlook with a wide range. Protected status: local protection. In the academic publication «Kazakhstan Flora» [6] D. maclata was combined with D. fuchsii (Druce) Soo. D. maculata hybridizes readily to form numerous transitional forms at both the interspecific and intergeneric levels [27, 28]. According to the works of M.G. Vakhrameeva [29, 30] D. maculata has a narrow ecological amplitude.
In the Kazakh Altai mountain ridge, in the eastern part of the Kalba ridge, small D. maculata populations were found. Coordinates of location: 49º 25´ N, 82º 36´ E, 791 m above sea level. In joint communities with D. incarnata various hybrid forms can be found. The species is confined to the marshy shaded edges of birch or aspen growths, or less often near willows. Rarely, individuals of the species are found in open areas of floodplain meadows, in communities with D. incarnata. The species is actively propagated by seed and vegetative means, by planting extra digitate tuber.
For primary introduction into the Altai Botanical Garden, it was introduced as living plants from the southern part of the Sibin depression of the Kalba ridge. In total, 35 generative individuals were planted in culture.
Sampled individuals D. maculata have a height of 28-36 cm. The stems are whole, without internal cavities, and are brittle and thickened at the base to 0.8-1 cm. The upper stem is indistinctly filled. The lower leaves of the sheath are quite loosely attached to the stem, while the upper ones are firmly attached and held close.
D. maculata has a cylindrical, clavate, or occasionally ovate inflorescence. Color variations of inflorescences can vary from lilac to dark purple. The sizes of the inflorescences are practically identical: length 44.5 cm, width 2.5 cm. In the case of bright saturated purple inflorescence, saturated round spots are always observed on the leaves. Flowers are normally developed, and no anomalies were detected. In one inflorescence there are 30-36 fully formed flowers. The lower floral lip is tripartite, and has a narrow middle part. The average leaf is rounded at the end or sometimes wedge-shaped, with a large number of longitudinal specks. The length of the lower lip is almost equal to its width. Side petals are of lanceolate form and slightly pointed. An elongated spur is located under the flower and has a cylindrical shape, 7-9 mm long. Wedgeshaped bracts prominently appear above inflorescence on lower flowers, upper bracts are almost equal in length to the flowers. The length of the lower bracts varies slightly from 15 to 16 (x = 15.6) mm, the width varies from 2 to 6 (x = 4.07) mm. All bracts are green with a weak purple border.

D. maculata is characterized by erect leaves, the lower of which may droop downwards. The shape of the majority of leaf blades is lanceolate, but there are also narrow-lanceolate leaves. A characteristic feature of the leaves in D. maculata is the presence of purple blotches. Spots are rather dull, sometimes imperceptible, and mostly small and dotted in form. They are arranged in horizontal stripes, sometimes resembling a transverse dotted line. The plant forms two bottom leaves or rarely one, which are always quite wide and drooping, and reach a length of 6.5–8 cm and a width of 2–3 (x = 2.3) cm. There are 2–4 erect leaves on the stem, from 8 to 10 (x = 9.3) cm in length and 0.7 to 2 (x = 1.24) cm wide. Not more than two bracts, always shortened, linear, and without violet border, from 2 cm in length and 3–6 (x = 4) mm in width.
The species is strongly susceptible with rust fungi.
Dactylorhiza umbrosa (aKar. & Kir.) Nevski (Fig. 6) is a rare meso-hygrophytic species from the Caucasian-Central Asian region [1]. Rarity category: 3b or R (b) — a rare species with a wide range, often growing in specific habitats. Protection status: local protection. It is a highly polymorphic species [31]. As a rule, plants are spread sparsely, in small groups or growing alone. The species does not form dense populations. In ecological terms, the species prefers humid places both in open, well-lit clearings, and in shade, such as under a forest canopy or shrub, in a valley of rivers or streams, or less often on flood meadows in motley grass. The soil substrate is usually represented by humus soils, with a large amount of overheating foliage of varying degrees of decomposition. In the phytocoenosis, the species steadily occupies territory, although it is often supplanted by sedges.
D. umbrosa was introduced into the Altai Botanical Garden collection as living plants from the eastern part of the Kalba ridge, the Koktau Mountains, from the valley of the river Taldybulak. The location coordinates: 49º 26´ N, 82º 33´ E, 696 m above sea level.
For planting samples of D. umbrosa in the exposition of natural flora, areas were selected that most closely corresponded to the natural ones: with constant shading under the canopy, richly humified and moistened soil with a significant layer of ground litter. Only 15 generative plants were introduced into the culture.
Dactylorhiza umbrosa specimens are powerful and multi-stemmed, with a large number of basal leaves. Plant height varies from 43 to 47 (x = 39.6) cm. There are 7–9 stems per individual, which are markedly curved, faceted at the top, and slightly thickened at the base to 7–10 (x = 8,5) mm. Stem shoots are collected in a fairly clear turf. Sods are well-defined, rounded, 8–18 (x = 13.7) cm in diameter. Color variations of inflorescences can vary from lilac-pink to dark pink shades. The inflorescences are straight, less curved, rather thick, 6–9 (x = 7.2) cm in length and 3.5–4 (x = 3.83) cm in width. The shape of the inflorescences is usually conical or cylindrical-cone-shaped, less often conical-ovoid. In one inflorescence there can be 46 to 64 (x = 52) fully formed flowers, without anomalies. The lower lip of the flower is tripartite, occasionally whole or solid-rhomboid. The color gradient is usually monochromatic, sometimes with a bright middle. The length of the lower lip is slightly larger than or equal to the middle. The middle part of the lower lip is almost always rounded, rather narrow, and often extends 1–2 mm forward. The lateral perianths are lanceolate or broadly lanceolate to oblong-rhombic, and slightly rounded at the ends. The spur of the flower is strongly elongated to 11–13 mm, is cylindrical in shape. The bracts are narrow, wedge-shaped, have a matte purple hue, and are always longer than the flowers, up to 15–18 (x =16.6) mm in length and 4–6 (x = 5) mm in width.
A characteristic morphological feature of this species is the abundant foliage of the stems and the presence of rosette basal leaves. The leaves are always present in large numbers, mostly drooping, lanceolate or

broadly lanceolate in form, slightly keeled, and light green in color. Young leaves are always keeled, lanceolate, and directed upward. Formed leaves have a wide groove along the entire length, are asymmetrical, markedly curved, and without spots, and are curved in the middle. There is not more than one lower basal leaf per stem shoot. As a rule, leaves of this type are creeping, drooping, and occasionally bent in the middle part, with a broadly lanceolate form, and are 8 to 15 (x =11 cm) in length, and 3 to 3.5 cm (x = 3,12 ± 0,3, Cv = 8 %) in width. There are 4-5 (x = 4.4) stem leaves, which rarely exceed the length of the stem, and are usually narrow lanceolate, loosely adjacent to the stem, and slightly deflected to the side or rarely directed upwards. The dimensions of the stem leaves vary from 11-25 (x = 16.2) cm in length and 1-2 (x = 1.6) cm in width. Bracts can have a violet hue or a brownish border around the edge of the leaf blade, are always linear-lanceolate in shape, and are 3.5-6 (x = 4.7) cm long and 6-10 cm (x = 7.8) cm wide. There are 1-2 bracts (x = 1.6 ± 0.3, Cv = 24 %) present, which are always well developed. Leaf sheathes are weakened and often friable, with lower leaves markedly elongated.
Conclusion
As a result of field expeditions in 2017 and 2018 on the Ivanov and Kalba ridges in the Kazakhstan part of Altai mountain system, 5 species of the genus Dactylorhiza were added to the Altai botanical garden for primary introduction by live plants: D. fuchsii (Druce) Soo, D. longifolia (L. Neum.) Aver., D. incarnata (L.) Soo, D. maculata (L.) Soo, and D. umbrosa (Kar. & Kir.) Nevski. These species in natural conditions of growth (in the Kazakh Altai mountain system) are found spontaneously, in separate individuals or in small groups. These species can be found along streams, on moss litter among the rocks, in flood meadows of wetland, and on the fringes of birch growth with admixture of aspen and willow with partial shading on the moist mixed grass meadows, both in the short grass and tall communities.
Thanks to the high level of habitat similarity in the area of introduction, particularly, the level of illumination, moisture and soil richness of nutrients, individuals showed a high percentage of rooting. The morphological characteristics and morphometric indicators of individuals of the species collected are the starting indicators when studying the introduction potential of the studied species of the genus Dactylorhiza.
The results obtained, as well as the collected samples of the genus Dactylorhiza, serve as the basis for molecular genetic analysis of species and intraspecific variability of Orchids, including a more accurate delineation of overlapping species using DNA technologies [32, 33].
This article was prepared within the framework of projects АР05133868 «Study of the distribution and current state of populations of species of the Orchid family of Kazakhstan Altai and their introduction in the Altai Botanical Garden» and AP05131621 «Information system on molecular genetics and botanical documentation of the wild flora of Kazakhstan» for 2018–2020.
References
- Vahrameeva, M.G., Varlygina, T.I., & Tatarenko, I.V. (2014). Orkhidnye Rossii (Biolohiia, ekolohiia i okhrana) [Orchid Russia (Biology, Ecology and Protection)]. Moscow: Tovarishchestvo nauchnykh izdanii KMK [in Russian].
- Mamaev, S.A., Knyazev, M.S., Kulikov, P.V., & Filippov, E.G. (2004). Orkhidnye Urala: sistematika, biolohiia, okhrana [Orchids of the Urals: systematics, biology, protection]. Ekaterinburg: UrO RAN [in Russian].
- Fardeeva, M.B., Bakin, O.V., & Izmailova, K.I. (2011). Struktura hibridnykh populiatsii Dactylorhiza maculata (L.) Soo S.L. na yuhe lesnoi zony [The structure of hybrid populations of Dactylorhiza maculata (L.) Soo S.L. in the south of the forest zone]. Proceedings from Protection and cultivation of orchids: IX Mezhdunarodnaia nauchnaia konferentsiia — IX International Scientific Conference (pp. 436—442). Moscow: Tovarishchestvo nauchnykh izdanii KMK [in Russian].
- Naczk, A.M., Chybicki, I, & Zietera, M.S. (2016). Genetic diversity of Dactylorhiza incarnata (Orchidaceae) in northern Poland. Acta Societatis Botanicorum Poloniae, 85(2): 3496, 1—14.
- Krasnaia kniha Altaiskoho kraia. Tom 1. Redkie i nahodiashchiesia pod uhrozoi ischeznoveniia vidy rastenii i hribov. (2016). [Red Book of the Altai Territory. Volume 1. Rare and endangered species of plants and fungi.]. Barnaul: Izdatelstvo Altaiskoho universiteta [in Russian].
- Pavlov, N.V. (Ed.). Flora Kazahstana [Flora of Kazakhstan]. (1958). (Vol. 2). Alma-Ata: Izd. AN KazSSR [in Russian].
- Krasnaia kniha Kazakhstana. Rasteniia [Red Book of Kazakhstan. Plants]. (2014). (Vol. 2; 2nd ed.). Astana [in Russian].
- Efimov, P.G. (2011). Orhidnye severo-zapada evropeiskoi chasti Rossii [Orchids of Northwest European part of Russia]. Moscow: Tovarishchestvo nauchnykh izdanii KMK [in Russian].
- Fedorov, A.L., Kirpichnikov, M.E., & Artyushenko, Z.T. (1956). Atlas po opisatelnoi morfolohii vysshikh rastenii. List. [Atlas on descriptive morphology of higher plants. Leaf]. Moscow: Izd. AN SSSR [in Russian].
- Fedorov, A.L., Kirpichnikov, M.E., & Artiushenko, Z.T. (1962). Atlas po opisatelnoi morfolohii vysshikh rastenii. Stebel i koren [Atlas on descriptive morphology of higher plants. Stem and root]. Moscow: Izd. AN SSSR [in Russian].
- Fedorov, A.L., & Artiushenko, Z.T. (1975). Atlas po opisatelnoi morfolohii vysshikh rastenii. Tsvetok. [Atlas on descriptive morphology of higher plants. Flower]. Leningrad: Nauka [in Russian].
- Fedorov, A.L., & Artiushenko, Z.T. (1979). Atlas po opisatelnoi morfolohii vysshikh rastenii. Sotsvetie [Atlas on descriptive morphology of higher plants. Inflorescence]. Leningrad: Nauka [in Russian].
- Bondartsev, A.S. (1954). Shkala tsvetov [Color scale]. Moscow: Izd. AN SSSR [in Russian].
- The color scale of the English Royal Society of Florists (RHS ColourChart).
- Zaitsev, G.N. (1990). Matematika v eksperimentalnoi botanike [Mathematics in Experimental Botany]. Moscow: Nauka [in Russian].
- Dospehov, B.A. (1985). Metodika polevoho opyta [Field experience]. Moscow: Ahropromizdat [in Russian].
- Cherepanov, S.K. (1995). Sosudistye rasteniia Rossii i sopredelnykh hosudarstv (v predelakh byvsheho SSSR) [Vascular plants of Russia and neighboring countries (within the former USSR)]. Saint Petersburg [in Russian].
- Almanah: Altaiskomu botanicheskomu sadu — 80 let. [Almanac: Altai Botanical Garden — 80 years]. (2015). Ust- Kamenogorsk: Izd. Media Alians [in Russian].
- Kamelin, R.V., Ovesnov, S.A., & Shilova, S.I. (1999). Nemoralnye elementy vo florakh Urala i Sibiri [Nemoral elements in the flora of the Urals and Siberia]. Perm: Izdatelstvo Permskoho universiteta [in Russian].
- Filippov, E.G., & Andronova, E.V. (2017). Heneticheskaia struktura populiatsii i estestvennaia hibridizatsiia Dactylorhiza salina i D. incarnata (Orchidaceae) [Genetic population structure and natural hybridization of Dactylorhiza salina and D. incarnata (Orchidaceae)]. Henetika — Genetics, 53, 3, 310-323 [in Russian].
- Ellenberg, H., Weber, H.E., Dull, R., Volkmar, W., Willg, W., & Dirk, P. (1991). Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica, 18, 1, 248.
- Vahrameeva, M.G., Tatarenko, I.V., & Bychenko, T.M. (1994). Ekolohicheskie harakteristiki nekotorykh vidov evraziatskikh orkhidnykh [Ecological characteristics of some species of Eurasian Orchid] Biulleten Moskovskoho obshchestva ispytatelei prirody. Otdel biolohicheskii — Bulletin of the Moscow Society of Naturalists. Biological department, 99, 4, 75-82 [in Russian].
- Sobko, V.G. (1989). Orkhidei Ukrainy [Orchids of Ukraine]. Kiev: Naukova dumka [in Russian].
- Alekhin, A.A. (1992). Predvaritelnye itohi introduktsii orkhidnykh flory SSSR v botanicheskom sadu Kharkovskoho universiteta [Preliminary results of the introduction of the orchid flora of the USSR in the Botanical Garden of Kharkov University]. Vestnik Kharkovskoho universiteta — Bulletin of Kharkov University, 364, 63-66 [in Russian].
- Efimov, P.G. (2012). Issledovaniia heneticheskoho polimorfizma Dactylorhiza baltica, D. fuchsii i D. incarnata (Orchidaceae) iz severo-zapada evropeiskoi chasti Rossii metodom ISSR [Studies of the genetic polymorphism of Dactylorhiza baltica, D. fuchsii and D. incarnata (Orchidaceae) from the north-west of the European part of Russia using the ISSR method]. Botanicheskii zhurnal — Botanical Journal, 97, 6, 751-760 [in Russian].
- Tsvelev, N.N. (2002). Opredelitel sosudistykh rastenii Severo-Zapadnoi Rossii (Leninhradskaia, Pskovskaia i Novhorodskaia oblasti) [Key to vascular plants of North-West Russia (Leningrad, Pskov and Novgorod regions)]. Saint Petersburg: Izdatelstvo SMPHFA [in Russian].
- Jagiello, M. (1990). Variability and distribution of some species from genus Dactylorhiza Necker ex Nevski in Poland. Acta Univ. Wratislawiensi, 1055, 45-55.
- Schmeidt, O. (1996). Estonia Orchideed. Tallinn: Varrak.
- Vakhrameeva, M.G., & Denisova, L.V. (1990). Nekotorye aspekty izucheniia redkikh vidov rastenii Moskovskoi oblasti (na primere orkhidnykh) [Some aspects of the study of rare plant species of the Moscow region (for example orchids)]. Ekolohiia i okhrana prirody Moskvy i Moskovskoho rehiona — Ecology and nature conservation in Moscow and the Moscow region (pp. 8487). Moscow: Izd. MGU [in Russian].
- Vakhrameeva, M.G. (2000). Rod Palchatokorennik [Genus Dactylorhiza]. Biolohicheskaia flora Moskovskoi oblasti — Biological flora of the Moscow region, 14. (pp. 55-86). Moscow: Grif i K [in Russian].
- Ovchinnikov, P.N. (Eds.). (1963). Flora Tadzhikskoi SSR. Sem. Orchidaceae [Flora of the Tajik SSR. Orchidaceae family]. Moscow; Leningrad: Izd. AN SSSR [in Russian].
- Turuspekov, Y., & Abugalieva, S. (2015). Plant DNA barcoding project in Kazakhstan. Genome, 58, 5. 290.
- Turuspekov, E.K., Ivashchenko, A.A., Ishmuratova, M.Yu., Kotuhov, Yu.A., Danilova, A.N., & Myrzagalieva, A.B., et al. (2017). Heneticheskoe raznoobrazie dikorastushchei flory Kazakhstana [Genetic biodiversity of Kazakhstan wild flora]. Proceedings from The study, conservation and rational use of the plant world of Eurasia, dedicated to the 85th anniversary of the Institute of Botany and Phytointegration: Mezhdunarodnaia nauchno-prakticheskaia konferentsiia — International scientific-practical conference. (pp. 143-148). Almaty [in Russian].