Бүйректң бұрылысты қарама-қарсы ағымды өзектік көбейту жүйегінде Генле iлмегiнщ ерекше конструкциясы қоюландыру қабiлеттiлiгiн жоғары дәрежеде ұстауды қамтамасыз етедi. Дегенмен, бүйректщ зақымдануын көптеген аурулар (диабет, гипертония, гломерулонефрит, интерстициалды нефрит ж.б.) шақыра алады. Олар алдында Yдерiске нефронның тек бiр сегментiн және онымен бiрге қан тамырларын, шумақтарды, өзектеpдi қатыстырады. Кейiннен Yдерiс нефронның кезкелген 6өлігін немесе оны қоршаған интерстициасын жазымдайды, нэтижесiнде әдетте бүйректщ көлемгі кiшiрейедi, бүйрек заңдылықты түрде қалыпты архитектоникасын жоғалтады.
Заманауи несеп түзу механизмі, несеп түзу теориясы - фильтрациялық - реабсорбциялық-секрециялық үдерісте жүретін осмостық сұйылту және осмостық қоюландыру бұрылысты қарама - қарсы ағымдағы өзекшелік көбейту желісті механизм. Нефрон өзекшелерінің барлық бөліктері милы қабаттың тамырлары, бүйректің интерстициалды тіні несепті осмостық қоюландыруына біріккен түрде қатысады. Демек, осы аталған бөліктер түгелдей - бұрылысты қарама- қарсы ағымдағы көбейту жүйесі функциясы ретінде бірыңғай, бірге жұмыс істейді.
Несеп шығару механизмдері, несеп түзу теориялары.
Бүйректің физиологиясын, патофизиологиясын ұғынуда, бүйрек физиологиясының тарихы, кімнің не істегені, нені анықтағаны бүйректің қисынын біліп тануды жеңілдетеді, жол ашады, үйретеді Карл Людвиг(1816-1895) және Гейденгаин (1834-1897) жазған, ұсынған пікірлер өзінің уақытында мағынасы маңызды болғанымен алынған мәліметтер көбінде жанама деректер сипатында болатын. ХХ ғасыр, несеп түзілу механизмі туралы деректерді алуда жаңа әдістемелерді, жаңа мүмкіндіктерді әкелді.
Мәселен, ағылшын дәрігері Ричардс нефрон капсуласын пункциялап, алынған «провизорлық» несептің құрамын «дефинитивті» (ақырғы) несептің құрамымен салыстырды. Бұл мәлімет өзекшелік диурездің мәнін анықтауға мүмкіндік берді. Американдық фармаколог А.Кешни, К.Людвигтің 1842 жылғы фильтрациялық теориясын қолдап өзінің «фильтрациялық-реабсорбциялық» несеп түзілу теориясын ұсынды. Соңғы кездерде бүйректі зерттеуде заманауи әдістемелерді, атап айтсақ изотоптық сынамаларды қолдану арқылы несеп түзілуінің механизмдері бүгінгі күндері қабылданған жаңа көзқарас жүзінде құрастырылды. Бұл жаңа теорияға Людвиг пен Гейденгайннің пікірлері де кірді.
ХХ ғасырдың орта шенінде Wirz H., Hargitay B., Kuhn W. (1951) несепті осмостық сұйылту және осмостық қоюландыру (концентрациялау) механизмі жөнінде идеяларын жариялап, оны эксперимент жүзінде дәлелдей білді. Бұл аса маңызды жаңалық нефрон шумақтарының бұрылысты қарама-қарсы ағымдықтағы өзекшелік көбейту жүйесі қызметінің үдерісіне негізделген. Бүгінгі таңда қалыптасқан пікір бойынша нефронның бөліктері, милы қабаттың қантамырлары, бүйректің интерстициальды тіні, әсіресе жинақтаушы түтікше, яғни, өзекшелердің барлығы түгелдей несепті осмостық қоюландыруына біріккен түрде бірегей барлығы жұмыла қатысады. Демек, осы аталған бөліктер түгелдей бұрылысты қарама-қарсы ағымдықтағы көбейту жүйесі функциясы ретінде бірыңғай, бірге жұмыс істейді.
Кейінгі кезде қабылданған теория: фильтрациялық - реабсорбциялық-секрециялық үдерісте жүретін осмостық сұйылту және осмостық қоюландыру, бұрылысты қарама- қарсы ағымдықтағы өзекшелік көбейту желісті механизм.
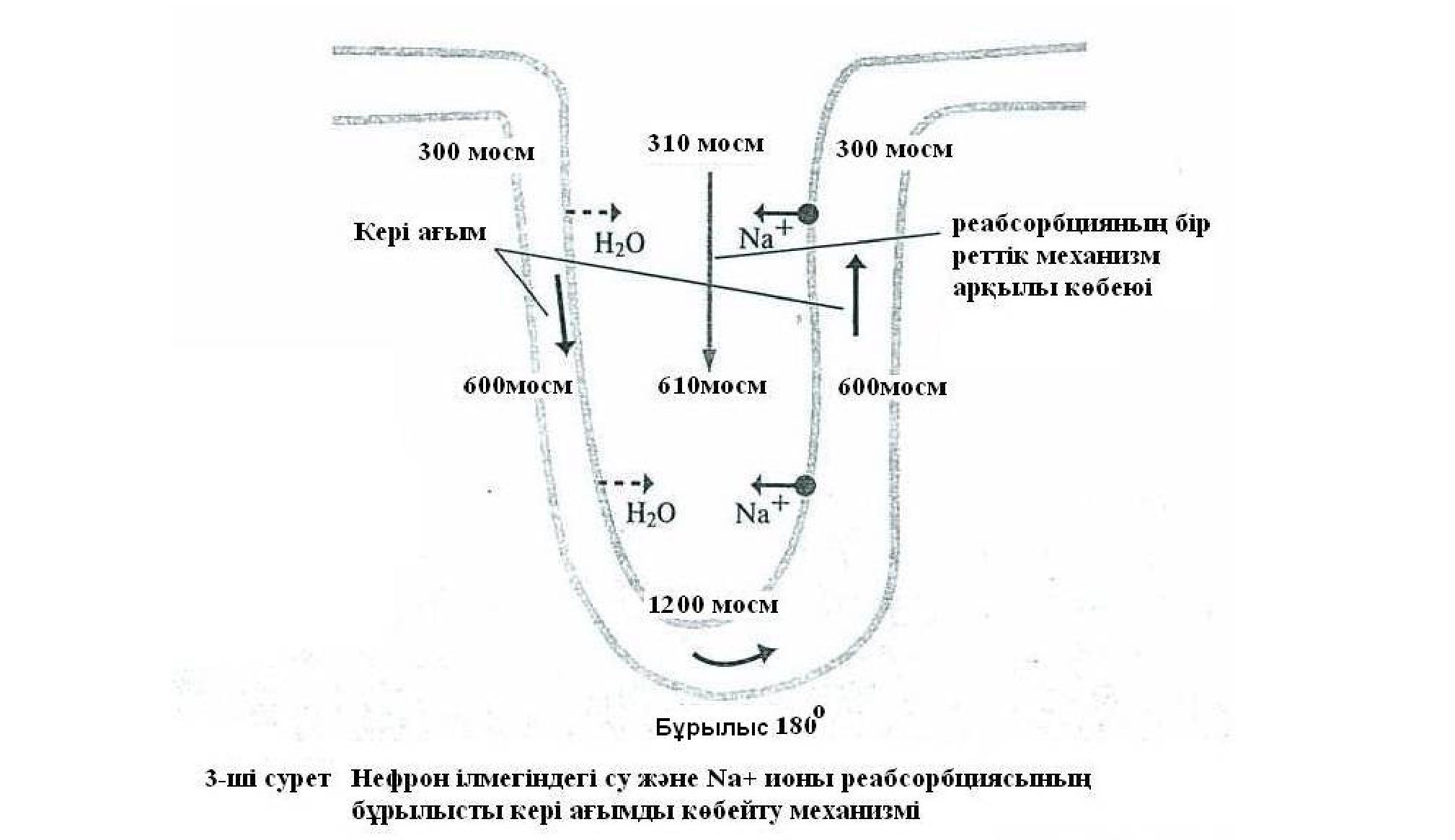
Осы теория аясында осмостық сұйылту және осмостық қоюландыру үдерісі, бұрылысты қарама-қарсы ағымдықтағы өзекшелік көбейту желісінің механизмі қалай істейді екен, қалай іске асады екен соған тоқталайық. Нефрон ілмектерінің әртүрлі бөліктерінің функционалдық мағынасы бір қатарлы, бірдей емес. Милы қабаттың сыртқы аумағындағы осмостық концентрацияның жоғары болуы Генле ілмегінің жоғары бағыттықтағы жуан бөлігінің қызметіне байланысты. Оның қабырғасы суды өткізбейді, ал жасушалары интерстициалды тінге натрийді, хлорды тасымалдайды. Ілмектің төменге бағытталған бөлігінің суға өткізгіштігі жоғары. Сондықтан, су өзекше саңлауларынан жан-жағын қоршаған интерстиция тініне осмостық градиент бойынша өтеді, ал осмос белсенді заттар өзекше қуысында қала береді. Милы қабаттың интерстициялық тініне хлор мен натрийдің түсуі бүйрек аумағының милы қабатында осмос белсенді заттардың жасушааралық сұйықтықтарда осмостық концентрациясының
жоғарылауына алып келеді. Генле ілмегінің төмен бағыттықтағы тік бөлігі мен иінінен судың интерстициялық тінге сорылуынан өзекше саңлауларында осы мезетте, сондай дәрежелікте осмостық концентрация өседі, арта түседі, жоғарылайды. Қыртысты қабатты заттардан бүйрек емізікшелерінің бойымен ілмектің төмен бағыттықтағы иінімен тереңдікте ұзаған сайын осмостық концентрция (қоюлануы) жоғарылай түседі. Нәтижесінде милы қабаттың бойымен төмен жылжығанда өзекше саңлауында және интерстиция тіндерінде біртіндеп осмостық концентрация (дені сау адамдарда) 290-нан 1200 мосмоль/кг дейін жетеді. Изотониялық 0,9% несеп гипертониялыққа 3,6% дейін жоғарылайды. Қалыптыда қан плазмасының осмостығы 287 мОсм/кг Н2О шамасында. Нефрон ілмегінің жоғары тіке бағытталған жуан иінінен натрий мен хлордың тінге реабсорбциясы жалғаса түседі, ал су өзекше саңлауында қала береді. Осы үдеріс бірте-бірте дисталды ирелеңді өзекшенің бастапқы бөлігіне жеткенде саңлаудағы сұйықтық барлық уақытта гипотониялық күйде болады, ал осмостық белсенді заттардың концентрациясы 200 мосмоль/кг аспайды. Дисталды ирелеңді өзекшеден сұйықтық (несеп) жинақтаушы түтікке түсердің алдында байланыстырушы өзекше арқылы өтеді. Осы жердегі бүйректің қыртысты қабатында (заттарында) су осмостық градиент бойынша сорылады (реабсорбцияланады), сұйықтықтарың концентрациясы 300 мосмоль/кг дейін көтеріледі. Әрі қарай жинақтаушы түтікшелердің милы қабатында несеп одан әрі жайлап, біртіндеп қоюлана түседі. Оның қоюлану дәрежесі милы қабаттың төменгі «шыңына» жеткенде интерстициядағы осмостық қысыммен пара - пар болады (1-ші және 2- сурет).
Ендеше, бүйректің несепті қоюландыруы немесе сұйылтуы бұрылысты қарама-қарсы ағымдықтағы өзекшелік көбейту жүйесі қызметі арқылы қамтамасыз етіледі, олар параллельді Генле ілмегі иіндері мен жинақтаушы түтікшелер құрылымдарымен бірге біріккен күйде қызмет етеді. Несеп өзекшелерде қарамақарсы бағытта қозғалыста болады (қарама-қарсы ағымдық деп аталуы содан), ал жүйенің бір иініндегі заттарды тасымалдау үдерісі екінші иіннің қызметі арқылы күшейе, көбейе («көбейеді») түседі. Төмен бағытталған иіннен, жоғары бағытталған иінге несеп өткенде 1800 бұрылыс жасайды. Нефрон ілмектерімен несеп жылжу барысында жеке реабсорбциялық механизмдер іске қосылып несеп құрамындағы заттар көбейтіледі, ол су және тұздардың санының айтарлықтай мол сорылуына ықпал етеді. Қарама-қарсы ағымдық механизмінде Генле ілмегінің жоғары бағытталған иіні (тік жуан иіні) бұл үдерісте шешуші ролді атқарады. Оның эпителийлері қоршаған интерстициалды кеңістікке натрий ионын белсенді түрде реабсорбциялайды да интерстициалды сұйықтың гиперосмостығын ұлғайтады, көтереді. Ендеше Генле ілмегі қоюландырушы (концентрациялаушы) механизм ретінде жұмыс істейді (1-ші және 2- сурет).
Натрийдің тасымалдануы калий және хлордың (Na-, К-, 2СL – *котранспортымен) және сутегі ионының **антипорт (айырбастау, обменник) атты тасушылардың көмегімен бірге қатарласа жүреді. Бұл жерде суға өткізгіштік өте төмен өйткені онда аквопориндер жоқ.
*Хлор анионымен ілеспелі түрде натрий ионы электрохимиялық градиент бойынша пассивті сорылады (реабсорбцияланады). Мұндай бір ионның (натрий) басқа заттың (хлорид) пассивті тасымалдануымен бірге жүруін котранспорт деп атайды.
** Өзекше эпителийінің апикалды мембранасында электронейтралды тасымалдаушы бар, ол Na+ және Н+ бірімен бірінің белсенді алмасуын қамтамасыз етеді, бұл кезде жасушадан кеткен Н ионының орнына натрий келіп орналасады, алмасады. Мұндай тасымалдау механизмі антипорт (алмасу, обменник) деп аталады.
Төмен қарай бағытталған ілмек иінінде аквопориннің 1-ші типі болғандықтан су пассивті (енжар) өзекше саңлауынан гиперосмостық интерстицияға өтеді. Олай болса, нефронның төменге бағытталған иінінде судың сорылуынан бара- бара гиперосмостық күйде болады, яғни интерстициалық сұйықтықпен осмостық тепе-теңдік (теңгерім) қалыптасады. Жоғары бағытталған иінде натрийдің сорылуына байланысты дисталды өзекшенің қыртысты бөлігінде несеп гипотониялық күйге жетеді. Бірақта, Генле ілмегінде су мен тұздардың сорылуынан несептің мөлшері айтарлықтай азаяды (1- сурет).
Содан кейін жинағыштық түтікке түскен несеп Генле ілмегінің жоғары бағыттағы иінімен қарама-қарсы ағымдық жүйе құрайды. Егерде жинағышты түтікшелерде су аз сорылса, ал натрий, хлоридтер, мочевинаның реабсорбциясы жалғаса түссе, онда нефрон несепті көп мөлшерде концентрациясы төмен гипотониялық күйде шығарады. Бұл несепті сұйылту үдерісі деп аталады. Ол ішкі ортадан судың артық мөлшерін шығару қажеттілігінде іске асады (бейімделулік, адаптациялық үдеріс). Несеп жинағыштық түтікше арқылы милы қабатқа тереңірек енген сайын су гиперосмостық интерстицияға пассивті түрде өтеді де несеп қоюлана түседі. Интерстиция сұйықтығындағы мочевина концентрациялық градиент бойынша Генле ілмегінің жоғары бағытты жіңішке иінінің саңлауына сорылады (диффузияланады) және де несеп ағымымен бірге дистальды өзекшелер мен жинағышты түтікшеге қайта түседі. Осылайша мочевинаның өзекшелердегі қайтымды, шеңберлі оралымды айналымы жүзеге асады, сөйтіп ол милы қабат концентрациясының жоғары деңгейлігін сақтайды, қамтамасыз етеді. Аталған мочевинаның тасымалдану үдерісі, негізінде, бүйректің милы қабатына терең бойлап түсуші Генле ілмегі ең ұзын юкстамедуллярлық нефрондарында жұмыс істейді.
Бүйректің милы
қабатында және де
қантамыр
капиллярларынан құралған қарама-қарсы ағымды қантамырлық жүйе бар. Юкстамедуллярлық нефрондардың қантамырлық торы өзекшелерге параллелді тік төмен және жоғары бағытты ұзын милы қабатқа терең бойлаушы капиллярлық тамырлар желісін құрайды. Қарама - қарсы ағымды тамырлық жүйе суды тартып алып кетуімен
бірге натрий және мочевинаны интерстицияда ұстауы арқылы бүйректің терең милы зат тіндерінде осмостық қысымның жоғары деңгейде болуын қамтамасыз етпейді, тек оның сақталуына болысады. Мочевинаның шеңберлік қайта ағымды айналымда болуы бүйрек нефронының ішкі милы затының осмостық қысымы жоғары болуына қажетті. Бүйректің бұл аймағында осмостық қысымның жоғары дәрежлігін екі фактор қамтамасыз етеді: (Na+ , СL) иондары және мочевина. Олардың бұған қосар үлесі де іс жүзінде бірдей десек болады.
ӘДЕБИЕТТЕР ТІЗІМІ
- Артур К. Гайтон, Джон Э. Холл. Медицинская физиология. - М.: Издательство «Логосфера», 2008. - 1256 с.
- Джеймс А. Шейман. Патофизиология почки. Перевод с англ., Под ред. акад., Ю.В.Наточина. - М.: Издательство БИНОМ, 2016. - 190 с.
- В.В.Новицкий. Патофизиология. - М.: «ГЭОТАР-Медиа», 2010. - 592 с.
- Ю.В.Наточин. Современный курс классической физиологии (Избранные лекции). - 2007. - 384 с.
- Чеснокова С.А. Карл Людвиг (1815-1895). - М.: «Наука», 1973. - 255 с.
- Чеснокова С.А. Рудольф Гейденгайн (1834-1897). - М.: Наука»,1978. - 144 с.